【大豆抗花叶病育种】
拼译:breeding of soybean resistant to soybean mosaic virus(SMV)
大豆花叶病为世界性大豆病害,流行广,危害重。抗大豆花叶病育种的目的是抵御大豆花叶病的发生和流行,控制病毒株系的消长,减免因品种感病造成的产量损失,降低因施用农药带来的环境污染及人体危害。
1899年,中国东北首次报道大豆花叶病。1916年,美国克林顿(G.P.Clinton)描述了大豆花叶病症状主要为花叶、卷叶和黄化等。1924年,加德纳(M.W.Gardner)等提出该病由危害株种子传播。1940年,德国海因策(K.Heinze)等证明该病也可由蚜虫传播。1948年,美国学者康诺弗(R.A.Conover)证明该病由一种以上的病毒株系引起。1954年,日本正式开展大豆花叶病研究,鉴别致病性不同的株系,鉴定来自本国及其他国家的品种抗性;1962年配制抗病组合;1977年推广了抗病品种出羽娘;1980年又推出一批新的抗病品种东北57、东北59等。美国的大豆花叶病不重,但抗病育种仍为主要育种目标之一。1977年,罗斯(J.P.Ross)通过回交育成抗、感大豆花叶病的近等基因系。1978年,伯纳德(R.L.Beranrd)采用回交方式将抗病品种PI96983的抗病基因导入优良品种威廉斯82(Williams82)中,提高了该品种的抗性。中国大豆抗花叶病育种工作开展的较晚,但南方早于北方。1975年,王国勋开始抗病育种的鉴定,1982年育成抗病品种中豆3、4和5号。林建兴利用齐黄1号、徐豆1号等为抗源,于1983年杂交育出科黄8号、科系8号和诱变30,是目前中国生产品种中抗性最强的品种。东北于1981年开始,1990年育成丰产、抗病品种吉林21号。韩国、巴西和阿根廷等国家也先后开展了抗大豆花叶病育种工作。例如1981年韩国推广的品种苏文97(Suweon97),已知的有7个株系免疫或高抗,并且丰产性好。1974年,朝鲜大豆花叶病毒株系——坏死株系的出现,使株系问题得到普遍重视。古德曼(R.M.Goodman)等利用一套抗性不同的大豆品种为鉴别寄主,将从美国品种资源上分离得到的98个毒株划分为7个株系:G1、G2、G3、G4、G5、G6、和G7。中国许志刚在此基础上增加一个鉴别寄主,将无法归入古德曼7株系的毒株定为G8、G9和G103个新株系。另外,日本高桥,中国濮祖芹、陈永宣、吕文清和罗瑞梧等也提出了自己的株系划分系统。但是,这些鉴定系统采用的鉴别寄主不同,局限性大,株系间吻合性不清楚,不便进行抗病品种的交流、引用。1984年,中国张明厚证明朝鲜的大豆花叶病坏死株系、吕文清的3号株系群为古德曼的G5。1979年,古德曼指出美国的标准毒株ATCCPV-94为他的G1;罗斯的疱疹株系为他的G3。目前,中国胡蕴珠和美国利姆(S.M.Lim)正在进行更加细致的工作。大豆花叶病的抗性由核基因控制。1982年,利姆(S.M.Lim)研究了苏文97对古德曼G2和G7株系的抗性遗传,从F1、F2和F3的分离结果发现该抗性由1对显性基因控制。多数研究结果支持这一结论。但也有不同结果,1980年,夸恩(S.H.Kwon)证明大豆对大豆花叶病坏死株系的抗性由一对隐性基因支配。1979年,罗梅乌(A.S.K.Romeu)等在品种PI96983中发现抗病基因,命名为Rsv1;在奥登(Odgen)中发现对Rsv1为隐性的等位基因,定名为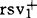 。两个等位基因对感病等位基因是显性。1984年,巴泽尔(R.I.Buzzell)等在雷电(Raiden)中发现的显性基因位于另一位点,定名为Rsv2。1989年,又在哥伦比亚(Columbia)品种中发现一个控制坏死反应的显性基因,位于第3位点,定名RSV3。1980年,鲍尔斯(G.R.Bowers)在品种哈迪(Hatdee)中发现一个对G5、G6和G7都有抗性,与抗源水牛(Buffalo)的抗性基因位点不同的显性基因,没有命名。1983年,鲁恩(G.W.Roane)研究证明抗大豆花叶病基因和抗花生花叶病基因间存在连锁,其重组率为3.7%±8%。1980年,希尔(J.H.Hill)等用小区罩网试验证明,大豆花叶病没有越冬寄主,带毒种子是病毒的初侵染源,蚜虫是大豆花叶病毒传播的主要介体。1979年,鲍尔斯等证明品种开花前接种种传率为18%,R1期接种种传率为11%,后期接种下降到4%。可见控制蚜虫,推迟品种感染时期,选择不传毒、不带毒或带毒率低的品种是必须的。1982年,布赖恩特(G.R.Bryant)建议建立种子检测制度,以利斯特(R.M.Listter)的酶联免疫吸附测验法应用最多。古德曼等已筛选出抗种传品种麦尔特(Merit)。1990年,中国杨书军等进行抗种传品种筛选指标研究,选出低种传品种浙春2号和夏豆82-16。种皮斑驳会影响大豆籽粒外观品质,降低大豆商品价值。1957年,日本越水幸男首次报道大豆花叶病可引致种皮斑驳。1981年,吕文清证明,低温、多雨利于感病植株褐斑粒形成。1991年,吴忠璞对大豆籽粒种皮斑驳发生规律的研究表明,花、荚期感病比苗期感病植株褐斑率高,抗斑驳可通过基因遗传。1991年,廖林研究证明,抗成株症状、抗种传和抗斑驳分别受不同基因控制,利用育种手段可选育出抗种传、抗斑驳和抗成株症状或兼抗三者的品种。1985年,曼里(Y.Maury)报道,不存在抗大豆花叶病毒所有株系的品种,对一个或几个株系高抗或免疫的材料也不多。古德曼对已鉴定的大豆品种再次筛选,获得高抗他的7个株系的品种苏文97。鲍尔斯从适于热带种植的大豆品系中筛选出抗古德曼7株系的品系PI424131。巴斯(G.R.Buss)认为,如果能获得目的基因,也会采用遗传工程或DNA重组技术进行抗病育种。1991年陈章良报道,利用基因工程已获得大豆抗花叶病的目的基因。继1974年周光宇提出外源DNA导入技术后,1986年刘得璞应用于大豆抗花叶病育种,1991年获得抗性得到改进的稳定品系。1985年,巴斯提出抗大豆花叶病为简单遗传,常规的育种方法即可快速、有效地转移抗性基因,回交或改良性回交最易凑效。但是,常规育种方法限制了抗病品种的选出频率,新的抗病品种必须具有低御新毒株的能力。因此,及时跟踪、鉴定大豆花叶病毒株系分类及消长动态,是育种家、植物病理学家和植保工作者共同关注的问题,也是抗病育种工作的首要问题。世界各大豆生产国都在开展抗大豆花叶病育种工作,随着品种抗性的提高同时表现出品质下降。如何获得抗病并且优质的品种,将是该领域很有研究价值的项目。研究现有各大豆花叶病毒株系划分系统的吻合性,明确抗源材料的抗性基因及遗传规律,结合生理、生化研究确定抗性基因位点及连锁图,开展抗性机理研究及抗性鉴定指标分析等,将是该领域最活跃的研究内容。另外,探索大豆花叶病毒毒力变化与抗病品种抗性丧失频率,也是一个很重要的课题。由于杂交品种的大面积推广,将导致遗传脆弱性,新毒株出现频繁,病虫害猖獗。所以,在拓宽抗源种质、培育抗地区优势毒株的品种的同时,开展抗多毒株育种;结合抗病毒病,开展抗孢囊线虫、灰斑病、霜霉病和细菌斑点病乃至一些虫害的多抗性育种或者多品系育种是该领域近期研究的中心课题。【参考文献】:1 Cho E K,Goodman R M.Phyto.,1979,69(5):4672 桥本钢二.农业かょび园艺,1980,7:23~283 Buss G R,et al.WSRC Ⅲ Proceeding,1986.433~4384 Ross J R,Virul and bacterial disease.Soybean:Improvement,production and uses,1987,2:728~7495 廖林,等.中国农学通报,1991,7(4):14~18
。两个等位基因对感病等位基因是显性。1984年,巴泽尔(R.I.Buzzell)等在雷电(Raiden)中发现的显性基因位于另一位点,定名为Rsv2。1989年,又在哥伦比亚(Columbia)品种中发现一个控制坏死反应的显性基因,位于第3位点,定名RSV3。1980年,鲍尔斯(G.R.Bowers)在品种哈迪(Hatdee)中发现一个对G5、G6和G7都有抗性,与抗源水牛(Buffalo)的抗性基因位点不同的显性基因,没有命名。1983年,鲁恩(G.W.Roane)研究证明抗大豆花叶病基因和抗花生花叶病基因间存在连锁,其重组率为3.7%±8%。1980年,希尔(J.H.Hill)等用小区罩网试验证明,大豆花叶病没有越冬寄主,带毒种子是病毒的初侵染源,蚜虫是大豆花叶病毒传播的主要介体。1979年,鲍尔斯等证明品种开花前接种种传率为18%,R1期接种种传率为11%,后期接种下降到4%。可见控制蚜虫,推迟品种感染时期,选择不传毒、不带毒或带毒率低的品种是必须的。1982年,布赖恩特(G.R.Bryant)建议建立种子检测制度,以利斯特(R.M.Listter)的酶联免疫吸附测验法应用最多。古德曼等已筛选出抗种传品种麦尔特(Merit)。1990年,中国杨书军等进行抗种传品种筛选指标研究,选出低种传品种浙春2号和夏豆82-16。种皮斑驳会影响大豆籽粒外观品质,降低大豆商品价值。1957年,日本越水幸男首次报道大豆花叶病可引致种皮斑驳。1981年,吕文清证明,低温、多雨利于感病植株褐斑粒形成。1991年,吴忠璞对大豆籽粒种皮斑驳发生规律的研究表明,花、荚期感病比苗期感病植株褐斑率高,抗斑驳可通过基因遗传。1991年,廖林研究证明,抗成株症状、抗种传和抗斑驳分别受不同基因控制,利用育种手段可选育出抗种传、抗斑驳和抗成株症状或兼抗三者的品种。1985年,曼里(Y.Maury)报道,不存在抗大豆花叶病毒所有株系的品种,对一个或几个株系高抗或免疫的材料也不多。古德曼对已鉴定的大豆品种再次筛选,获得高抗他的7个株系的品种苏文97。鲍尔斯从适于热带种植的大豆品系中筛选出抗古德曼7株系的品系PI424131。巴斯(G.R.Buss)认为,如果能获得目的基因,也会采用遗传工程或DNA重组技术进行抗病育种。1991年陈章良报道,利用基因工程已获得大豆抗花叶病的目的基因。继1974年周光宇提出外源DNA导入技术后,1986年刘得璞应用于大豆抗花叶病育种,1991年获得抗性得到改进的稳定品系。1985年,巴斯提出抗大豆花叶病为简单遗传,常规的育种方法即可快速、有效地转移抗性基因,回交或改良性回交最易凑效。但是,常规育种方法限制了抗病品种的选出频率,新的抗病品种必须具有低御新毒株的能力。因此,及时跟踪、鉴定大豆花叶病毒株系分类及消长动态,是育种家、植物病理学家和植保工作者共同关注的问题,也是抗病育种工作的首要问题。世界各大豆生产国都在开展抗大豆花叶病育种工作,随着品种抗性的提高同时表现出品质下降。如何获得抗病并且优质的品种,将是该领域很有研究价值的项目。研究现有各大豆花叶病毒株系划分系统的吻合性,明确抗源材料的抗性基因及遗传规律,结合生理、生化研究确定抗性基因位点及连锁图,开展抗性机理研究及抗性鉴定指标分析等,将是该领域最活跃的研究内容。另外,探索大豆花叶病毒毒力变化与抗病品种抗性丧失频率,也是一个很重要的课题。由于杂交品种的大面积推广,将导致遗传脆弱性,新毒株出现频繁,病虫害猖獗。所以,在拓宽抗源种质、培育抗地区优势毒株的品种的同时,开展抗多毒株育种;结合抗病毒病,开展抗孢囊线虫、灰斑病、霜霉病和细菌斑点病乃至一些虫害的多抗性育种或者多品系育种是该领域近期研究的中心课题。【参考文献】:1 Cho E K,Goodman R M.Phyto.,1979,69(5):4672 桥本钢二.农业かょび园艺,1980,7:23~283 Buss G R,et al.WSRC Ⅲ Proceeding,1986.433~4384 Ross J R,Virul and bacterial disease.Soybean:Improvement,production and uses,1987,2:728~7495 廖林,等.中国农学通报,1991,7(4):14~18(吉林省农业科学院廖林副研究员撰)