【DNA复制】
拼译:DNA replication
DNA储存有生物体的遗传信息,研究DNA复制就是探索生物体增殖的规律。DNA复制研究的内容包括DNA复制的方式、起点、方向和速度;DNA合成的引发;DNA链的延伸;DNA合成的真实性和进行性;DNA合成的终止和DNA链的分离,以及DNA子链中错误的修正等。许多酶和蛋白质参与DNA复制,包括DNA聚合酶及附属蛋白、解链酶、旋转酶、拓扑异构酶、单链DNA结合蛋白、引发酶、DNA连接酶,以及与DNA复制起始有关的蛋白等。分离纯化这些酶和蛋白质,并在体外重建出有完全活性的复制复合体是DNA复制研究的主要目标之一。研究DNA复制的起始,研究参与DNA复制的酶和蛋白质的基因表达的调控及蛋白质功能的修饰调节,对于了解生物体增殖规律具有重要意义。
自从20世纪50年代中科恩伯格(A.Kornberg)等首先从大肠杆菌分离纯化出DNA聚合酶Ⅰ以来,原核生物的DNA复制研究已经取得很大进展。对DNA合成过程与DNA聚合酶作用已研究得比较清楚,而对DNA复制的调控还知道得不多,对真核生物的DNA复制研究则差距较大。通常DNA合成的开始要有一段RNA链作引物,合成这段引物的酶称引发酶,各种生物的引发酶差别很大。真核的引发酶与DNA聚合酶α紧密结合在一起;原核则不同,有的引发酶与解链酶结合在一起(T4噬菌体),有的甚至与解链酶在一条多肽链中(T7噬菌体)。大肠杆菌中引发酶与dnaB、dnaC、dnaT等近20条多肽链一起组成一个多功能的引发体。不同的引发酶作用机制差别也很大。有的识别几个碱基,有的合成引物较长但与其顺序无关,有的只以核糖核苷酸作底物,有的也以脱氧核糖核苷酸为底物合成RNA-DNA-RNA这样一种交替的引物。现在还不清楚引发酶如何识别它的起动子以及引物的合成如何终止。另外,所有引发酶都从5’→3’合成引物。在DNA合成时后随链的合成方向与复制叉前进的方向是相反的。引发酶如何在总体上随着复制叉沿着模板链的5’→3’方向运动,而又以模板链的3’→5’为依据合成引物的机制也不很清楚。除了以RNA作引物外,也有一些例外。一些噬菌体与质粒靠专一的核酸内切酶水解产生3’OH作为DNA合成的引物。有的单链病毒像Parvoviruses靠3’末端折叠配对作为引物。腺病毒和枯草杆菌噬菌体ф29则依靠专一的蛋白与核酸结合作为引物。现在有些病毒、微生物和动物细胞的引发酶的基因已分离到,正在进一步进行研究。在DNA新链的延伸中,DNA聚合酶表现出极高的真实性和非常高的合成效率。DNA合成的真实性由两步来达到。首先,DNA聚合酶的聚合活性能选择正确的脱氧核糖核苷酸,参入到引物的3’末端,这也称“碱基选择”。这一步的出错几率约为10-4~10-5。第二步由DNA聚合酶的3’→5’的外切活性切去已参入到引物3’末端的错误的核苷酸,称为“校正阅读”。这一步可使总的出错几率再下降两个数量级,使总出错率达到10-6以下。DNA合成的真实性,尤其对于DNA聚合酶在“碱基选择”中的作用机制,仍是目前研究的热点之一。原核生物DNA聚合酶大多具有3’→5’外切活性,真核生物DNA聚合酶δ和ε具有3’→5’外切活性。说明由3’→5’的校正阅读来保持DNA复制真实性的机制在原核和真核中也是相同的。DNA聚合酶的催化效率取决于DNA聚合酶催化反应的进行性。如果酶在将一个核苷酸加在DNA链上以后从引物末端解离下来再结合上去则需要约1min,而连续进行合成则只需1左右。已分离得到DNA聚合酶有的反应进行性高,有的则较低。酶反应进行性高低与它们在体内的功能有关,例如大肠杆菌DNA聚合酶Ⅰ进行性低,它的功能是填补DNA链的空缺,在体内主要在修复DNA时起作用。大肠杆菌DNA聚合酶Ⅲ合成进行性高,主要在复制DNA时起作用。一些多亚基的DNA聚合酶的进行性经常与其中的某一亚基有关,如DNA聚合酶Ⅲ中的τ亚基及DNA聚合酶δ的附属蛋白。T7噬菌体则利用大肠杆菌的硫氧还蛋白作为亚基来使DNA聚合酶具有高的进行性。DNA聚合酶的催化合成反应的进行性仍然是目前研究的题目之一。对DNA聚合酶的结构和构象现在还知道得很少,现在唯一测出三维结构的是大肠杆菌DNA聚合菌I的C端大片段,分辨率为2.75×10-10m。从这个三维模型看,酶的聚合活性和3’→5’外切活性位于两个功能区中,酶与DNA复合物的构象以及酶在催化DNA合成时构象的变化仍是需要解决的问题。现用X-衍射已测到DNA聚合酶I大片段与DNA处于外切状态复合物的三维结构。用扫描隧道显微镜(STM)也观察到DNA聚合酶Ⅰ大片段与DNA复合物的一种构象。分离纯化DNA聚合酶的各亚基及其它蛋白因子,研究它们的功能,再重建出具有完全活性的DNA复制体,才能说是对某一生物担负DNA复制的酶系统的一个完整的研究结果。目前,大肠杆菌DNA聚合酶Ⅲ基本上可以说达到了上述目标。真核生物DNA聚合酶和其它复制蛋白的研究从80年代中以来取得了突破性的进展,斯蒂尔曼(B.Stillman)实验室等用猿猴病毒40(SV40)建立的体外DNA复制体系提供了有力的工具。从SV40环状双链DNA上的复制起始点开始进行的DNA合成,除需要由病毒基因编码的T抗原外,其他则利用宿主细胞的酶和复制因子。利用这一体系,证明DNA聚合酶δ及分裂细胞核抗原(PCNA)是SV40进行完整而有效的复制所必需的,从而确定除DNA聚合酶α外,DNA聚合酶δ也是细胞DNA的复制酶。用杂交方法证明,DNA聚合酶δ合成前导链,DNA聚合酶α合成前导链的第一条冈崎片段以及后随链。用SV40体外体系还分离纯化了RF-C等复制因子。DNA聚合酶反应的物理化学研究得还较少。酶如何与单链的、双链的DNA结合,如何运动到引物末端,酶与DNA作用各状态的能级及构象,复制起始蛋白或复制体与复制起始点的识别还不太清楚。研究得较多的还是大肠杆菌DNA聚合酶Ⅰ的大片段,用快速动力学研究了聚合过程的基本步骤,测定酶上二价金属离子结合位点时发现在底物dNTP及NMP存在时各有一个二价金属离子结合点由弱变强。据测定,引物3’末端结合到酶的3’→5’外切活性时至少要拆开4对碱基。对大肠杆菌DNA聚合酶Ⅲ的研究表明,它依靠ATP的活化与模板引物相结合。利用ATP类似物研究证明,有两种性质有差别的DNA聚合酶Ⅲ。利用SV40体外体系已证明在引物末端从DNA聚合酶α向DNA聚合酶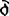 转换的机制,在这一转换中,RF-C和PCNA起到识别引物末端及帮助转换的作用。DNA复制叉的前进,要依靠解链酶来打开DNA双链,还要由拓扑异构酶来松弛超螺旋,单链结合蛋白可保护解开的DNA单链不被核酸酶水解,又防止它们重新回复成双链。正是这些众多的蛋白质帮助完成了DNA复制过程。根据大肠杆菌DNA复制提出来的假说认为,在复制叉的部位复制体有两个极相似的DNA聚合酶,它们随着复制叉前进方向一起前进,一个聚合酶合成前导链,另一个聚合酶合成后随链;后随链合成时通过成环使DNA链的延伸方向与复制叉的前进方向保持一致。当合成一定长度后环解开,又在近复制叉处形成新环。真核生物与原核生物有些差别,复制前导链与后随链的两个酶是不同的酶,而原核中两个酶的催化亚基是相同的。真核中已鉴定参与DNA复制的蛋白质还有单链结合蛋白RF-A、PRP1和PRP2两种引物识别蛋白,DNA聚合酶α/引物酶的活化因子AAF、解链酶、拓扑异构酶。染色体DNA复制如何起始是一个很不清楚的问题。大肠杆菌DNA复制起始点oriC是个245碱基对的区域。大肠杆菌的带有oriC区域的质粒给研究提供了便利。现发现有许多蛋白参与大肠杆菌DNA复制的起始,包括dnaA、dnaB、dnaC、HU蛋白、RNA聚合酶、拓扑异构酶I和RNaseH。引发酶、DNA聚合酶、解链酶、旋转酶和单链结合蛋白也参与DNA复制的起始。dnaA蛋白与oriC位点的相互作用使oriC区的DNA双链打开,在其它蛋白的共同作用下开始合成DNA。细菌内dnaA蛋白含量对DNA复制起始有很大影响,影响超螺旋密度的蛋白质(拓扑异构酶,旋转酶,HU)以及复制起始区附近的转录等因素也会影响DNA复制的起始。噬菌体DNA复制起始有它们自己的调控机制。对与真核生物基因复制的起始,现在也利用病毒DNA作模型来进行研究。Papovavirus情况与大肠杆菌相似。SV40则有T抗原结合到专一的起始区域,用它的解链酶活性打开双链并开始复制。真核细胞基因组比原核大得多,分别安排在几十条染色体上,为确保DNA的有效复制,每条染色体上多点起始进行复制。在酵母中已鉴定出自复制顺序(ARS),带ARS的质粒能在酵母中自发复制,也已鉴定出与ARS结合的蛋白质。DNA复制如何在ARS上起始还需进一步研究。人染色体DNA复制起始也是多点起始且顺序专一性不高。此外,真核染色体DNA在一个细胞周期只能复制一次的机制还不清楚。总之,关于染色体DNA复制起始仍有许多工作要做。已发现线状染色体DNA端点具有重复顺序,端区的存在与染色体的稳定及细胞分裂有关。在DNA合成结束后,由端聚酶在端区加长重复顺序。端聚酶是蛋白质和RNA的复合物,其中RNA是合成端区重复顺序的模板,蛋白质则是一种反转录酶。DNA子链中的错误将由DNA修复系统来进行修正。DNA修复系统将依据DNA链上甲基化程度的高低来区别母链与子链。对于这些过程及作用机制还需要进一步研究。原核生物两条DNA双链的分开据信与膜的结合有关。真核生物则依据微管的作用,染色体通过着丝粒与微管相连,在有丝分裂时沿着微管朝纺锤体端点移动。对参与DNA复制的蛋白质的基因的克隆的研究也取得不少进展。如原核生物DNA聚合酶Ⅲ的各个亚基已克隆,DNA聚合酶Ⅰ的克隆为研究它的作用机理创造了条件。近年来真核DNA聚合酶及其他复制因子的基因研究也取得不少进展。例如,人DNA聚合酶α,δ,酵母DNA聚合酶Ⅰ、Ⅱ、Ⅲ,人、鼠、果蝇等的PCNA均已克隆成功。比较它们的核苷酸顺序发现,人DNA聚合酶α与酵母DNA聚合酶Ⅰ,DNA聚合酶δ与酵母DNA聚合酶Ⅲ同源性很高,证明从低等真核生物酵母到哺乳动物DNA复制的蛋白质和机制都是保守的。现已对真核DAN聚合酶重新命名,使之统一起来。虽然DNA复制研究已取得不少进展,但仍有许多问题尚不清楚。例如对大肠杆菌DNA聚合酶Ⅱ、真核DNA聚合酶ε的功能还不清楚。复制蛋白质的修饰(如磷酸化)与它们功能的关系也未搞清。由于DNA复制与其他学科(例如细胞生物学,病毒学,肿瘤研究等)有着密切的关系.DNA复制研究的进展将为这些学科的发展创造条件。【参考文献】:1 Blow J. Nature, 1987,326:441~442 Kornberg A,J Biol Chem, 1988,263:1~ 42 Mcttenry C S. Annu Rev Biochem. 1988.57:519~5503 Beese L S,and steity T A. Nucleic Acids and Molecular Biology ed. by Eckstein. F. Liltey , D M .1 (Spring - vcrlag Berlin Heidelberg), 1989,3:28~434 Burgers P M J. Prog Nucleic Acid Res. 1989.37:235 ~ 2805 Tsurimoto,T Melendy, T. stillman B. Nature. 1990, 346: 534 ~5396 So A G,Downey K M. Critical Reviews m Biochemistry and Molecular Biolosy, 1992 , 27 : 129 ~ 155
转换的机制,在这一转换中,RF-C和PCNA起到识别引物末端及帮助转换的作用。DNA复制叉的前进,要依靠解链酶来打开DNA双链,还要由拓扑异构酶来松弛超螺旋,单链结合蛋白可保护解开的DNA单链不被核酸酶水解,又防止它们重新回复成双链。正是这些众多的蛋白质帮助完成了DNA复制过程。根据大肠杆菌DNA复制提出来的假说认为,在复制叉的部位复制体有两个极相似的DNA聚合酶,它们随着复制叉前进方向一起前进,一个聚合酶合成前导链,另一个聚合酶合成后随链;后随链合成时通过成环使DNA链的延伸方向与复制叉的前进方向保持一致。当合成一定长度后环解开,又在近复制叉处形成新环。真核生物与原核生物有些差别,复制前导链与后随链的两个酶是不同的酶,而原核中两个酶的催化亚基是相同的。真核中已鉴定参与DNA复制的蛋白质还有单链结合蛋白RF-A、PRP1和PRP2两种引物识别蛋白,DNA聚合酶α/引物酶的活化因子AAF、解链酶、拓扑异构酶。染色体DNA复制如何起始是一个很不清楚的问题。大肠杆菌DNA复制起始点oriC是个245碱基对的区域。大肠杆菌的带有oriC区域的质粒给研究提供了便利。现发现有许多蛋白参与大肠杆菌DNA复制的起始,包括dnaA、dnaB、dnaC、HU蛋白、RNA聚合酶、拓扑异构酶I和RNaseH。引发酶、DNA聚合酶、解链酶、旋转酶和单链结合蛋白也参与DNA复制的起始。dnaA蛋白与oriC位点的相互作用使oriC区的DNA双链打开,在其它蛋白的共同作用下开始合成DNA。细菌内dnaA蛋白含量对DNA复制起始有很大影响,影响超螺旋密度的蛋白质(拓扑异构酶,旋转酶,HU)以及复制起始区附近的转录等因素也会影响DNA复制的起始。噬菌体DNA复制起始有它们自己的调控机制。对与真核生物基因复制的起始,现在也利用病毒DNA作模型来进行研究。Papovavirus情况与大肠杆菌相似。SV40则有T抗原结合到专一的起始区域,用它的解链酶活性打开双链并开始复制。真核细胞基因组比原核大得多,分别安排在几十条染色体上,为确保DNA的有效复制,每条染色体上多点起始进行复制。在酵母中已鉴定出自复制顺序(ARS),带ARS的质粒能在酵母中自发复制,也已鉴定出与ARS结合的蛋白质。DNA复制如何在ARS上起始还需进一步研究。人染色体DNA复制起始也是多点起始且顺序专一性不高。此外,真核染色体DNA在一个细胞周期只能复制一次的机制还不清楚。总之,关于染色体DNA复制起始仍有许多工作要做。已发现线状染色体DNA端点具有重复顺序,端区的存在与染色体的稳定及细胞分裂有关。在DNA合成结束后,由端聚酶在端区加长重复顺序。端聚酶是蛋白质和RNA的复合物,其中RNA是合成端区重复顺序的模板,蛋白质则是一种反转录酶。DNA子链中的错误将由DNA修复系统来进行修正。DNA修复系统将依据DNA链上甲基化程度的高低来区别母链与子链。对于这些过程及作用机制还需要进一步研究。原核生物两条DNA双链的分开据信与膜的结合有关。真核生物则依据微管的作用,染色体通过着丝粒与微管相连,在有丝分裂时沿着微管朝纺锤体端点移动。对参与DNA复制的蛋白质的基因的克隆的研究也取得不少进展。如原核生物DNA聚合酶Ⅲ的各个亚基已克隆,DNA聚合酶Ⅰ的克隆为研究它的作用机理创造了条件。近年来真核DNA聚合酶及其他复制因子的基因研究也取得不少进展。例如,人DNA聚合酶α,δ,酵母DNA聚合酶Ⅰ、Ⅱ、Ⅲ,人、鼠、果蝇等的PCNA均已克隆成功。比较它们的核苷酸顺序发现,人DNA聚合酶α与酵母DNA聚合酶Ⅰ,DNA聚合酶δ与酵母DNA聚合酶Ⅲ同源性很高,证明从低等真核生物酵母到哺乳动物DNA复制的蛋白质和机制都是保守的。现已对真核DAN聚合酶重新命名,使之统一起来。虽然DNA复制研究已取得不少进展,但仍有许多问题尚不清楚。例如对大肠杆菌DNA聚合酶Ⅱ、真核DNA聚合酶ε的功能还不清楚。复制蛋白质的修饰(如磷酸化)与它们功能的关系也未搞清。由于DNA复制与其他学科(例如细胞生物学,病毒学,肿瘤研究等)有着密切的关系.DNA复制研究的进展将为这些学科的发展创造条件。【参考文献】:1 Blow J. Nature, 1987,326:441~442 Kornberg A,J Biol Chem, 1988,263:1~ 42 Mcttenry C S. Annu Rev Biochem. 1988.57:519~5503 Beese L S,and steity T A. Nucleic Acids and Molecular Biology ed. by Eckstein. F. Liltey , D M .1 (Spring - vcrlag Berlin Heidelberg), 1989,3:28~434 Burgers P M J. Prog Nucleic Acid Res. 1989.37:235 ~ 2805 Tsurimoto,T Melendy, T. stillman B. Nature. 1990, 346: 534 ~5396 So A G,Downey K M. Critical Reviews m Biochemistry and Molecular Biolosy, 1992 , 27 : 129 ~ 155(中国科学院上海生物化学研究所陆长德副研究员撰)