【光呼吸作用及其生化特征】
拼译:photorespiration and its biochemical character
植物的绿色组织在光下吸收氧气和释放二氧化碳的生理过程,称光呼吸。这个过程只有在光下才能进行。现知光呼吸与光合作用密切相关,只有在光合作用进行时才能发生光呼吸。
1968年美国Tolbert提出光呼吸概念。但是在此之前,人们已知在完整叶片中CO2的同化产物有乙醇酸、甘氨酸、丝氨酸等化合物的存在。至1971年他提出氧化光合碳循环的整个化学历程,以后有所补充。光呼吸是一个氧化过程。被氧化的底物是乙醇酸。乙醇酸是光合作用产物之一,它主要是通过1,5-二磷酸核酮糖羧化酶-加氧酶(RubisCO酶)作用生成的。20世纪70年代发现该酶具有双重催化功能之后,对光呼吸的底物乙醇酸来自光合作用才得到肯定的。RubisCO酶既可催化1,5-二磷酸核酮糖(RuBP)在Mg2+参与下羧化成二分子的磷酸甘油酸,也可以催化RuBP在Mg2+参与下加氧生成一分子磷酸甘油酸与一分子磷酸乙醇酸。磷酸乙醇酸在磷酸酯酶催化下转变为乙醇酸。这就是光呼吸底物的来源。此酶在催化羧化反应时称RuBP羧化酶,催化加氧反应时则称RuBP加氧酶。具体催化反应如下:



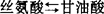 )组成。整个循环的总反应是:2磷酸乙醇酸+谷氨酸+O2+NADH+H++ATP+H2O→磷酸甘油酸+α·酮戊二酸+ADP+2Pi+NAD++NH3+CO2+2H+从上式可以看出,每2分子磷酸乙醇酸转变为一分子磷酸甘油酸即放出一分子的CO2。由光合作用所同化的碳,因光呼吸循环会损失掉1/4。整个循环涉及叶绿体、过氧体与线粒体3个细胞器。要维持光呼吸的循环正常运行,还原当量(NADH)的正常供应是必需的。1971年,Tolbert提出氧化光合碳循环的整个化学历程时,即提出由苹果酸与天冬氨酸穿梭作用来完成的设想。1983年,Schmitt等人用黑麦完整的过氧体实验,认为此穿梭系统不能提供过氧体以NADH。原因是:(1)产生NADH的苹果酸脱氢酸和谷草转氨酸的活性不足以维持光呼吸循环的速率。(2)尽管过氧体内苹果酸同功酶占细胞总量的40%,但谷草转氨酶只占细胞的不到7%。(3)加入苹果酸和谷氨酸后,过氧体只能缓慢地还原加入的NAD,在加入外源的谷草转氨酶后,NAD+的还原速率才会极大地增高。(4)供应苹果酸和谷氨酸,在缺少外源的NAD时,过氧体不能将羟基丙酮酸还原成甘油酸,只有加入外源NAD,这种还原反应才能进行。1984年,梁峥等用菠菜完整过氧体实验证实,由羟基丙酮酸→丝氨酸的转变中,最大活性不是在加入谷氨酸时获得,而是在加入丙氨酸后获得的;他们还发现,由甘油酸转变为丝氨酸过程中,NAD、丙氨酸和甘油酸是绝对必需的,并可用α酮戊二酸和天冬氨酸代替草酰乙酸。1987年,梁峥对过氧体内甘油酸转变为丝氨酸时NAD的供应做了实验,结果证实过氧体膜能缓慢地透过NAD/NADH,提出膜的转运载体可能参与菠菜叶片过氧体NAD/NADH的再产生的穿梭系统。由于在整个乙醇酸循环中,每二分子的乙醇酸释放出一分子的CO2,余下一分子磷酸甘油酸再入还原光合碳循环。即在一般大气条件下,光呼吸氧化的碳元素要占光合固定碳元素的25%~30%。由于光呼吸不断进行,光合强度下降可达50%,使净光合效率降低,因此关于光呼吸这一植物正常生命活动是有利还是有害,迄今仍未澄清,也是国内外研究者急于解决的课题。从能量代谢方面来看,光合作用的光合磷酸化为CO2的同化提供了ATP和NADPH,并将光能量转变为同化产物中的键能。呼吸作用则分解有机物释放其中的键能生成ATP和NAD(P)H,以便用于细胞内各种生命活动。光呼吸乙醇酸循环生成的中间产物如甘氨酸、丝氨酸可供合成蛋白质及其它化合物的原料,而磷酸甘油酸进入C3循环补充了循环的碳架,推动C3的循环,此是合成过程的一部分,是与光合作用分不开的。甘氨酸氧化中发生脱羧反应,释放出CO2招致碳的损失,但伴随着脱氨反应与NADH的生成又为谷氨酸的合成与ATP的生成做了准备,所以乙醇酸循环既合成又分解,既氧化又还原,既吸能又放能,这是与其它循环不同之处。1983年,陈锦强等用高水平硝态氮培育黄麻植株,其光合强度、光呼吸光合比值及硝酸还原酶活性、乙醇酸氧化酶活性均比铵盐为氮源培养的植株高,用抑制甘氨酸脱羧反应抑制剂异烟肼处理叶圆片,则硝酸还原受阻,说明光呼吸与硝酸还原具有一定联系,可能是由草酰乙酸-苹果酸穿梭为在细胞质使硝酸还原提供了还原剂NADH,如果乙醇酸循环中甘氨酸脱羧反应产生的NADH确能为硝酸还原提供还原剂的话,光呼吸在硝酸盐的同化中确会起积极作用。乙醇酸循环中,甘氨酸的脱羧是在线粒体内进行的,以NAD+为其电子受体。在线粒体内三羧酸循环(TCA)中的苹果酸脱氢酶与异柠檬酸脱氢酶也是以NAD+作为电子受体。80年代初,一些研究者已开始研究线粒体内甘氨酸氧化与TCA循环的关系,试图说明光呼吸与暗呼吸之间的关系。1990年,张少红用豌豆叶片线粒体的实验证实,线粒体中甘氨酸、苹果酸与异柠檬酸的氧化速率均受光的促进。甘氨酸氧化会抑制苹果酸与异柠檬酸的氧化,而其本身不受影响。用异烟肼抑制剂处理,或是提高NAD+的浓度均会降低甘氨酸氧化时对苹果酸氧化与异柠檬酸氧化的抑制。线粒体氧化甘氨酸、苹果酸与异柠檬酸的Km(NAD+)值分别是66.67μmol/L、119.1μmol/L和152.2μmol/L。甘氨酸氧化时抑制苹果酸与异柠檬酸的氧化是由于甘氨酸在竞争NAD+中占了优势。已知绿色叶片在照光下TCA循环仍在进行,以提供合成过程所需要的碳架,但TCA循环在光下运行较暗中慢,TCA循环的限速酶-异柠檬酸脱氢酶的活性受抑制是其主要原因。还有不少研究者从光、温度等方面做了研究。已知在饱和光强以上照射植株叶片常会引起光合抑制,此种光合抑制现象不但在叶片,就是在叶绿体与类囊体也能观察到。此种现象在碳代谢受限制的条件下尤为明显。其实质是多余的光能不能排散而引起光氧化。1978年,,Powles等用离体叶片在无CO2与不同O2浓度下进行光合抑制处理,发现随着O2浓度从7%提高到21%,光合抑制迅速被缓解,甚至完全消除。因而认为O2促光呼吸起着保护作用。已知绿色叶片经照光产生的Mehler反应是一个不可避免的过程,并藉此排除多余的能量。显然,植株是通过光呼吸与Mehler反应以保护光合作用的正常进行。1989年,王荣福等人分别用两种水稻品种进行实验。他们发现在1%~30%的O2浓度范围内,无CO2,在2000μE·M-2·S-1光照条件下,处理3h,两种水稻光合抑制程度会随着处理期间O2浓度的增高而被缓解。但是当O2浓度高于30%,两种水稻均会出现光抑光合加重现象,并且将这样处理的样品放置在正常生长条件与800μE·M-2·S-1时,光合速率分别被抑制28%与20%。他们认为光呼吸在光抑光合中只能在短时间内起缓解作用,O2通过Mehler反应来排除多余的能量,以及超氧物歧化酶(SOD)清除由Mehler反应产生的活性氧(O2、OH、O2、H2O2),保护光合膜的作用可能性则更为主要。植株在逆境(大气污染、低温、干旱、盐渍、强光)的条件下,体内的活性氧代谢会失调,在一定程度上会增加机体内活性氧的含量,从而加剧膜脂过氧化作用,破坏生物膜的结构,使膜差别透性丧失,代谢紊乱,植株衰老并遭受伤害。现知植株在逆境条件下呼吸强度会发生不同程度的改变。光呼吸作用是否可以看作是植物进行光合作用的一种抗逆性的表现,关键是植株叶片细胞内的乙醇酸氧化酶(GO)与SOD之间关系是怎样的。1991年,杨礼锐等用不同浓度的Na2SO3处理菠菜叶片,得知GO活性的变化与SOD活性的变化相一致,呈平行关系,而且低浓度的处理中,GO活性的改变是SOD活性改变的2.5-3倍,当
)组成。整个循环的总反应是:2磷酸乙醇酸+谷氨酸+O2+NADH+H++ATP+H2O→磷酸甘油酸+α·酮戊二酸+ADP+2Pi+NAD++NH3+CO2+2H+从上式可以看出,每2分子磷酸乙醇酸转变为一分子磷酸甘油酸即放出一分子的CO2。由光合作用所同化的碳,因光呼吸循环会损失掉1/4。整个循环涉及叶绿体、过氧体与线粒体3个细胞器。要维持光呼吸的循环正常运行,还原当量(NADH)的正常供应是必需的。1971年,Tolbert提出氧化光合碳循环的整个化学历程时,即提出由苹果酸与天冬氨酸穿梭作用来完成的设想。1983年,Schmitt等人用黑麦完整的过氧体实验,认为此穿梭系统不能提供过氧体以NADH。原因是:(1)产生NADH的苹果酸脱氢酸和谷草转氨酸的活性不足以维持光呼吸循环的速率。(2)尽管过氧体内苹果酸同功酶占细胞总量的40%,但谷草转氨酶只占细胞的不到7%。(3)加入苹果酸和谷氨酸后,过氧体只能缓慢地还原加入的NAD,在加入外源的谷草转氨酶后,NAD+的还原速率才会极大地增高。(4)供应苹果酸和谷氨酸,在缺少外源的NAD时,过氧体不能将羟基丙酮酸还原成甘油酸,只有加入外源NAD,这种还原反应才能进行。1984年,梁峥等用菠菜完整过氧体实验证实,由羟基丙酮酸→丝氨酸的转变中,最大活性不是在加入谷氨酸时获得,而是在加入丙氨酸后获得的;他们还发现,由甘油酸转变为丝氨酸过程中,NAD、丙氨酸和甘油酸是绝对必需的,并可用α酮戊二酸和天冬氨酸代替草酰乙酸。1987年,梁峥对过氧体内甘油酸转变为丝氨酸时NAD的供应做了实验,结果证实过氧体膜能缓慢地透过NAD/NADH,提出膜的转运载体可能参与菠菜叶片过氧体NAD/NADH的再产生的穿梭系统。由于在整个乙醇酸循环中,每二分子的乙醇酸释放出一分子的CO2,余下一分子磷酸甘油酸再入还原光合碳循环。即在一般大气条件下,光呼吸氧化的碳元素要占光合固定碳元素的25%~30%。由于光呼吸不断进行,光合强度下降可达50%,使净光合效率降低,因此关于光呼吸这一植物正常生命活动是有利还是有害,迄今仍未澄清,也是国内外研究者急于解决的课题。从能量代谢方面来看,光合作用的光合磷酸化为CO2的同化提供了ATP和NADPH,并将光能量转变为同化产物中的键能。呼吸作用则分解有机物释放其中的键能生成ATP和NAD(P)H,以便用于细胞内各种生命活动。光呼吸乙醇酸循环生成的中间产物如甘氨酸、丝氨酸可供合成蛋白质及其它化合物的原料,而磷酸甘油酸进入C3循环补充了循环的碳架,推动C3的循环,此是合成过程的一部分,是与光合作用分不开的。甘氨酸氧化中发生脱羧反应,释放出CO2招致碳的损失,但伴随着脱氨反应与NADH的生成又为谷氨酸的合成与ATP的生成做了准备,所以乙醇酸循环既合成又分解,既氧化又还原,既吸能又放能,这是与其它循环不同之处。1983年,陈锦强等用高水平硝态氮培育黄麻植株,其光合强度、光呼吸光合比值及硝酸还原酶活性、乙醇酸氧化酶活性均比铵盐为氮源培养的植株高,用抑制甘氨酸脱羧反应抑制剂异烟肼处理叶圆片,则硝酸还原受阻,说明光呼吸与硝酸还原具有一定联系,可能是由草酰乙酸-苹果酸穿梭为在细胞质使硝酸还原提供了还原剂NADH,如果乙醇酸循环中甘氨酸脱羧反应产生的NADH确能为硝酸还原提供还原剂的话,光呼吸在硝酸盐的同化中确会起积极作用。乙醇酸循环中,甘氨酸的脱羧是在线粒体内进行的,以NAD+为其电子受体。在线粒体内三羧酸循环(TCA)中的苹果酸脱氢酶与异柠檬酸脱氢酶也是以NAD+作为电子受体。80年代初,一些研究者已开始研究线粒体内甘氨酸氧化与TCA循环的关系,试图说明光呼吸与暗呼吸之间的关系。1990年,张少红用豌豆叶片线粒体的实验证实,线粒体中甘氨酸、苹果酸与异柠檬酸的氧化速率均受光的促进。甘氨酸氧化会抑制苹果酸与异柠檬酸的氧化,而其本身不受影响。用异烟肼抑制剂处理,或是提高NAD+的浓度均会降低甘氨酸氧化时对苹果酸氧化与异柠檬酸氧化的抑制。线粒体氧化甘氨酸、苹果酸与异柠檬酸的Km(NAD+)值分别是66.67μmol/L、119.1μmol/L和152.2μmol/L。甘氨酸氧化时抑制苹果酸与异柠檬酸的氧化是由于甘氨酸在竞争NAD+中占了优势。已知绿色叶片在照光下TCA循环仍在进行,以提供合成过程所需要的碳架,但TCA循环在光下运行较暗中慢,TCA循环的限速酶-异柠檬酸脱氢酶的活性受抑制是其主要原因。还有不少研究者从光、温度等方面做了研究。已知在饱和光强以上照射植株叶片常会引起光合抑制,此种光合抑制现象不但在叶片,就是在叶绿体与类囊体也能观察到。此种现象在碳代谢受限制的条件下尤为明显。其实质是多余的光能不能排散而引起光氧化。1978年,,Powles等用离体叶片在无CO2与不同O2浓度下进行光合抑制处理,发现随着O2浓度从7%提高到21%,光合抑制迅速被缓解,甚至完全消除。因而认为O2促光呼吸起着保护作用。已知绿色叶片经照光产生的Mehler反应是一个不可避免的过程,并藉此排除多余的能量。显然,植株是通过光呼吸与Mehler反应以保护光合作用的正常进行。1989年,王荣福等人分别用两种水稻品种进行实验。他们发现在1%~30%的O2浓度范围内,无CO2,在2000μE·M-2·S-1光照条件下,处理3h,两种水稻光合抑制程度会随着处理期间O2浓度的增高而被缓解。但是当O2浓度高于30%,两种水稻均会出现光抑光合加重现象,并且将这样处理的样品放置在正常生长条件与800μE·M-2·S-1时,光合速率分别被抑制28%与20%。他们认为光呼吸在光抑光合中只能在短时间内起缓解作用,O2通过Mehler反应来排除多余的能量,以及超氧物歧化酶(SOD)清除由Mehler反应产生的活性氧(O2、OH、O2、H2O2),保护光合膜的作用可能性则更为主要。植株在逆境(大气污染、低温、干旱、盐渍、强光)的条件下,体内的活性氧代谢会失调,在一定程度上会增加机体内活性氧的含量,从而加剧膜脂过氧化作用,破坏生物膜的结构,使膜差别透性丧失,代谢紊乱,植株衰老并遭受伤害。现知植株在逆境条件下呼吸强度会发生不同程度的改变。光呼吸作用是否可以看作是植物进行光合作用的一种抗逆性的表现,关键是植株叶片细胞内的乙醇酸氧化酶(GO)与SOD之间关系是怎样的。1991年,杨礼锐等用不同浓度的Na2SO3处理菠菜叶片,得知GO活性的变化与SOD活性的变化相一致,呈平行关系,而且低浓度的处理中,GO活性的改变是SOD活性改变的2.5-3倍,当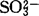 浓度超出一定阈值(100ppm)后,GO活性下降也较SOD活性下降的少,预示GO对环境改变较之SOD更为敏感。可能环境因子首先作用于GO,然后包括SOD在内的保护酶系统再行使各自的生化反应。1988年,Sandalio等人在西瓜的过氧体中发现有SOD。1989年,Rio等人见到豌豆叶片过氧体膜上NADH诱导出超氧自由基,提出
浓度超出一定阈值(100ppm)后,GO活性下降也较SOD活性下降的少,预示GO对环境改变较之SOD更为敏感。可能环境因子首先作用于GO,然后包括SOD在内的保护酶系统再行使各自的生化反应。1988年,Sandalio等人在西瓜的过氧体中发现有SOD。1989年,Rio等人见到豌豆叶片过氧体膜上NADH诱导出超氧自由基,提出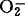 的产生可能是过氧体的普遍代谢性质,并进一步推论过氧体膜上存在有与活性氧有关的作用。有关RuBP羧化酶活性与乙醇酸氧化酶活性之间的关系,是1982年高煜珠等人用不同水稻品种间的实验获得的。他们证实凡是RuBP羧化酶活性较高的品种,其乙醇酸氧化酶的活性一般也较高,因此它们之间的相互关系也从生化基础上反映了光合作用与光呼吸之间的相互关系。随着分子生物学的发展,人们已着手对RuBP羧化酶的基因进行改造。主要是改造RuBP羧化酶的大亚基,加强其羧化能力削弱其加氧作用,从而加强光合强度降低光呼吸强度,以达到增产目的。另一方面,则是从改造乙醇酸氧化酶的基因入手。1986年Gerdes等人报道,扁豆转绿时总RNA和mRNA的含量增加与乙醇酸氧化酶的活性有平行关系。用分离编码的乙醇酸氧化酶的mRNA体外转译合成乙醇酸氧化酶蛋白,与从植物提取的乙醇酸氧化酶分子量(43kd)一致。随着与光呼吸相关的酶的mRNA分离的成功,编码这些酶的基因将被克隆,定会使人们对光呼吸机理的认识更向前推进一步。【参考文献】:1 Chang C C,et al.Plant Physiol.1981,67(5):1003~10062 Schmitt M R,et al.Plant Physiol.1983,72(3)728~7433 李明启.植物学通报,1985,2(1):1~74 梁峥.云南植物研究,1987,9(4):449~4545 王荣福,等.江苏农业科学,1988,4(2):1~86 Sandalio L M,et al.Plant Physiol.1988,88(4):1215~12187 del Rio,L A,et al.Plant Physiol.1989,89(3):728~7318 张少红,等.植物生理学报,1990,16(2):185~1918 杨礼锐,等.植物学通报,1991,81(1):43~47
的产生可能是过氧体的普遍代谢性质,并进一步推论过氧体膜上存在有与活性氧有关的作用。有关RuBP羧化酶活性与乙醇酸氧化酶活性之间的关系,是1982年高煜珠等人用不同水稻品种间的实验获得的。他们证实凡是RuBP羧化酶活性较高的品种,其乙醇酸氧化酶的活性一般也较高,因此它们之间的相互关系也从生化基础上反映了光合作用与光呼吸之间的相互关系。随着分子生物学的发展,人们已着手对RuBP羧化酶的基因进行改造。主要是改造RuBP羧化酶的大亚基,加强其羧化能力削弱其加氧作用,从而加强光合强度降低光呼吸强度,以达到增产目的。另一方面,则是从改造乙醇酸氧化酶的基因入手。1986年Gerdes等人报道,扁豆转绿时总RNA和mRNA的含量增加与乙醇酸氧化酶的活性有平行关系。用分离编码的乙醇酸氧化酶的mRNA体外转译合成乙醇酸氧化酶蛋白,与从植物提取的乙醇酸氧化酶分子量(43kd)一致。随着与光呼吸相关的酶的mRNA分离的成功,编码这些酶的基因将被克隆,定会使人们对光呼吸机理的认识更向前推进一步。【参考文献】:1 Chang C C,et al.Plant Physiol.1981,67(5):1003~10062 Schmitt M R,et al.Plant Physiol.1983,72(3)728~7433 李明启.植物学通报,1985,2(1):1~74 梁峥.云南植物研究,1987,9(4):449~4545 王荣福,等.江苏农业科学,1988,4(2):1~86 Sandalio L M,et al.Plant Physiol.1988,88(4):1215~12187 del Rio,L A,et al.Plant Physiol.1989,89(3):728~7318 张少红,等.植物生理学报,1990,16(2):185~1918 杨礼锐,等.植物学通报,1991,81(1):43~47(华南农业大学杨礼锐副教授撰)