【黄籽油菜】
拼译:yellow-seeded oilseed rape
黄籽油菜属十字花科芸苔属,其种皮较薄,种子含油量和蛋白质含量均较黑籽油菜的种子高,饼粕中纤维素含量较黑籽的低,因而极大地改善了菜籽油和饼粕的商品品质。培育黄籽油菜已成为油菜育种的重要目标之一。
黄籽油菜首先是由印度柏菜因(Prain)报道。1898年,他详细描述了在印度旁遮普省种植的白菜型油菜黄沙逊的黄籽性状。后来有人在印度的另一个白菜型油菜变种类型托利亚中发现了黄籽类型。因而有关白菜型黄籽油菜的形态、生理及遗传研究首先在印度开展起来,并引起各国育种家的重视。芥菜型黄籽油菜的研究由中国学者孙逢吉(1945)提出。1963年,阿桑那(N.A.Asthana)找到了芥菜型油菜黄皮品系“东方芥”。1968年,耐加(G.G.Nagar)报道了芥菜型油菜的黄种皮突变体。加拿大从“东方芥”中育成黄籽品种“多毛”。甘蓝型油菜黄籽的研究,于1960年由瑞典沃尔逊(Olsson)最先报道,以后在加拿大、德国均有过报道,但研究工作未见实质性进展,直至1979年刘后利从甘/白杂种后代获得甘蓝型黄籽后,甘蓝型黄籽油菜研究才受到各国育种家的重视。黄籽油菜种皮薄,种皮比率为12%~14%,而黑籽油菜的种皮比率为16%~19%。斯均根姆(Stringam,1974)对油菜种皮解剖结构研究结果是,黄籽种皮栅栏层细胞较黑籽薄得多。高永同于1981年测定,华油3号黑籽的种皮厚度为83.07u,而黄籽种皮厚仅为48.29u。肖达人(1982)证实,种皮色泽与种皮厚度呈正相关(r=0.9766)。相同遗传背景、不同种皮色种子含油量分析结果,黄籽含油量比黑籽增加1.54%~4.26%。1976年,斯坦利(Stanley)对芜菁油菜种子化学成分分析结果,粗纤维主要集中在种皮中,黄籽油菜的粗纤维含量为10.25%,黑籽为13.0%左右。有人认为通过黄籽育种可将油菜种子的粗纤维含量下降40%。遗传研究表明,油菜种皮色受母本植株基因型的控制并遗传。黄籽对黑(褐)籽为隐性,正反交种子种皮色均表现为母本种皮色,F1种子为深色(黑色、褐色),F2种皮色产生分离。油菜种皮色主要受核基因控制遗传,与细胞质基因无关。1942年,穆罕默德(A.Mohammad)对4个不同种皮色(黑褐色、红褐色、黄褐色、黄色)的白菜型油菜品系间的杂种F2研究结果,认为有3种基因控制白菜型油菜种皮色遗传,即Br1、Br2和Br3。Br1控制黑褐色种皮、Br2控制红褐色种皮、Br3控制黄褐色种皮的形成,而且Br1br1、Br2br2对Br3br3有上位性。只有3个位点基因均处于纯合隐性(br1br1br2br2br3br3)时才表现为黄籽。1980年,斯均根姆(G.R.Strirgam)用黄沙逊与3个加拿大白菜型油菜杂交,结果表明种皮色受两对独立的显性基因(Br1和Br3)控制。仅Br1位点为纯合隐性时,种皮表现为黄褐色,只有当两个位点处于纯合隐性时种皮才表现为黄色。1982年,斯切威卡(A.Schwetka)对7个白菜型黄籽油菜品系间双列杂交试验研究结果,认为油菜种皮色的基因包括2个上位性位点和4个下位性位点。黄籽的等位基因有两个,一个是位于Br1br2位点上的brbr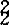 ,另一个是位于Br6Br6位点上的br
,另一个是位于Br6Br6位点上的br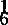 。白菜型油菜种脐颜色受一独立基因(Br8Br8)控制。在Br6Br6位点上存在着一个控制淡色种脐的基因(br
。白菜型油菜种脐颜色受一独立基因(Br8Br8)控制。在Br6Br6位点上存在着一个控制淡色种脐的基因(br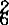 )。1942年,孙逢吉研究结果,芥菜型油菜种皮色受两对重叠基因(R1R2)控制,这一结果于1979年得到瓦拉(C.Vera)、1983年,埃斯拉姆(A.M.Aslam)等研究者的证实。1979年,刘后利报道甘蓝型油菜种皮色受3对以上基因控制。1986年,悉依亚得根(M.Shirz-adegam)对4个黄籽品系与一个黑籽品系正反交F2种皮色分离,结果证实甘蓝型油菜种皮色受3对基因(BL1BL2BL3)控制,当3个位点均为隐性时种皮为黄色。自然资源中,甘蓝型油菜无黄籽品种存在。目前的甘蓝型油菜黄籽品系主要来自甘蓝型油菜与白菜型黄籽油菜的种间杂交后代。甘蓝型黄籽油菜的种皮色显著不同于白菜型的和芥菜型的油菜,表现为土黄色或姜黄色。研究资料表明,黄籽性状的表达受环境条件影响较大。黄籽品种随种子成熟度的提高,种皮色逐渐加深。黄籽材料种皮色的变异是客观存在的。同一植株不同结角部位的种子种皮色表现不一致,年度间种皮色差异较大。1988年刘后利发现,甘蓝型黄籽油菜虽经长期选择、自交纯化,始终不能获得纯黄籽品系。黄籽油菜存在着种皮遗传不稳定性。这种不稳定性可能源自甘蓝的“CC”染色体组上黑种皮基因对白菜的“AA”染色体组上黄种皮基因的上位性;另一方面甘蓝型油菜种皮色变异的原因除与基因互作有关外,也可能与AC两个染色体组间的互作及环境条件影响有关。1987年,王汉中通过对大量甘蓝型黄籽品系种皮特性的比较研究发现,黄籽种皮表面分布着不均等的黑色斑点、斑块及环状斑,这些特征不存在于白菜型的和芥菜型的黄籽油菜种皮上,认为甘蓝型油菜黄籽性状可能受转座因子控制。黄籽种皮转色经由绿→黄绿→黄→黄褐→褐→黑褐的过程。这一过程表明,黄籽性状表达先于黑籽性状表达。1977年,辛达(Theander)等对芜菁油菜的生化分析结果证实,黄籽品系的多酚类化合物含量要比黑籽品系低得多,认为存在于种皮栅栏层中的多酚类化合物的积累量直接影响种皮色泽。1988年,胡晓君研究发现,黑籽油菜种子中花色素含量明显高于黄籽的,并鉴定出黑籽中含有的花色素类型主要为矢车菊素、锦葵素和飞燕草素等。1991年,王汉中应用组织化学方法检定了油菜种子皮壳中的多酚氧化酶活性的存在部位与粒色关系,结果表明白菜型的、甘蓝型的油菜在种子成熟前皮壳栅栏层中多酚氧化酶活性较强。黄籽皮壳栅栏层多酚氧化酶活性略低于黑籽的,但当有足够的底物存在时,黄籽种皮色也会逐渐转黑。而用抗坏血酸抑制多酚氧化酶活性后,种子保持黄色,说明多酚氧化酶对油菜转色有重要影响。由于甘蓝型黄籽油菜遗传基础的复杂性以及遗传不稳定性,在现有甘蓝型品种中进行杂交选育纯黄种子比较困难。1983年,埃斯拉姆提出用甘蓝型黄籽的与埃塞俄比亚芥黄籽的品种杂交,进行“CC”染色体组代换,以培育纯黄种皮甘蓝型油菜。1991年,戚存扣通过对该组合杂种F1染色体减数分裂行为观察结果,认为将埃芥“CC”染色体组上的黄籽基因转移到甘蓝型油菜中是可能的。因为在杂种F1代,来自埃芥的和甘蓝型油菜的两组“CC”染色体组的染色体能完全配对。1987年,陈宝元提出用淡黄籽的甘蓝品种与含黄沙逊黄种皮基因的白菜型品种杂交,人工合成甘蓝型黄籽材料。黄籽油菜,尤其是甘蓝型黄籽油菜,将是今后油菜育种研究的主要课题之一。目前,有关油菜种皮色的形成机制还很不清楚,需要运用现代分析技术研究油菜种皮中的生理生化过程,以求发现与种皮色直接相关的化学物质或生理生化过程,为油菜种皮色的遗传控制提供基础。中国栽培的白菜型的、芥菜型的油菜中,存在大量的黄籽农家品种,对这些品种的收集、整理,将丰富黄籽油菜研究的内容。有关甘蓝型油菜种皮色遗传研究,还有许多问题有待解决,如为什么在甘蓝型油菜中不能获得纯黄种子?粒色遗传不稳定性的原因是什么?应用现代生物技术和遗传工程手段进行基因组代换,分子原位杂交,将外源种质的黄种皮基因转移到甘蓝型油菜中来,将是未来黄籽油菜育种研究的一个重要方面。【参考文献】:1 Vera C L,et al.Can Plant Scl.,1979,59:635~6372 Stringam G R.Can Plant Sci.,1980,60:331~3353 SchwetKa A.Theor Appl Genet,1982,62:161~1694 肖达人.作物学报,1983,8(4):245~2545 高永同.中国油料,1984,3:85~896 Shirzadegam M,G Robbelen.FETTE.SEIFEN,1985,6:235~2377 刘后利.作物杂志,1988,2:5~78 傅寿仲,唐继宏,戚存扣.江苏农业学报,1988,4(1):26~309 戚存扣,仲裕泉,等.江苏农业学报,1991,7(4):8~1210 王汉中.中国油料,1991,1:30~33
)。1942年,孙逢吉研究结果,芥菜型油菜种皮色受两对重叠基因(R1R2)控制,这一结果于1979年得到瓦拉(C.Vera)、1983年,埃斯拉姆(A.M.Aslam)等研究者的证实。1979年,刘后利报道甘蓝型油菜种皮色受3对以上基因控制。1986年,悉依亚得根(M.Shirz-adegam)对4个黄籽品系与一个黑籽品系正反交F2种皮色分离,结果证实甘蓝型油菜种皮色受3对基因(BL1BL2BL3)控制,当3个位点均为隐性时种皮为黄色。自然资源中,甘蓝型油菜无黄籽品种存在。目前的甘蓝型油菜黄籽品系主要来自甘蓝型油菜与白菜型黄籽油菜的种间杂交后代。甘蓝型黄籽油菜的种皮色显著不同于白菜型的和芥菜型的油菜,表现为土黄色或姜黄色。研究资料表明,黄籽性状的表达受环境条件影响较大。黄籽品种随种子成熟度的提高,种皮色逐渐加深。黄籽材料种皮色的变异是客观存在的。同一植株不同结角部位的种子种皮色表现不一致,年度间种皮色差异较大。1988年刘后利发现,甘蓝型黄籽油菜虽经长期选择、自交纯化,始终不能获得纯黄籽品系。黄籽油菜存在着种皮遗传不稳定性。这种不稳定性可能源自甘蓝的“CC”染色体组上黑种皮基因对白菜的“AA”染色体组上黄种皮基因的上位性;另一方面甘蓝型油菜种皮色变异的原因除与基因互作有关外,也可能与AC两个染色体组间的互作及环境条件影响有关。1987年,王汉中通过对大量甘蓝型黄籽品系种皮特性的比较研究发现,黄籽种皮表面分布着不均等的黑色斑点、斑块及环状斑,这些特征不存在于白菜型的和芥菜型的黄籽油菜种皮上,认为甘蓝型油菜黄籽性状可能受转座因子控制。黄籽种皮转色经由绿→黄绿→黄→黄褐→褐→黑褐的过程。这一过程表明,黄籽性状表达先于黑籽性状表达。1977年,辛达(Theander)等对芜菁油菜的生化分析结果证实,黄籽品系的多酚类化合物含量要比黑籽品系低得多,认为存在于种皮栅栏层中的多酚类化合物的积累量直接影响种皮色泽。1988年,胡晓君研究发现,黑籽油菜种子中花色素含量明显高于黄籽的,并鉴定出黑籽中含有的花色素类型主要为矢车菊素、锦葵素和飞燕草素等。1991年,王汉中应用组织化学方法检定了油菜种子皮壳中的多酚氧化酶活性的存在部位与粒色关系,结果表明白菜型的、甘蓝型的油菜在种子成熟前皮壳栅栏层中多酚氧化酶活性较强。黄籽皮壳栅栏层多酚氧化酶活性略低于黑籽的,但当有足够的底物存在时,黄籽种皮色也会逐渐转黑。而用抗坏血酸抑制多酚氧化酶活性后,种子保持黄色,说明多酚氧化酶对油菜转色有重要影响。由于甘蓝型黄籽油菜遗传基础的复杂性以及遗传不稳定性,在现有甘蓝型品种中进行杂交选育纯黄种子比较困难。1983年,埃斯拉姆提出用甘蓝型黄籽的与埃塞俄比亚芥黄籽的品种杂交,进行“CC”染色体组代换,以培育纯黄种皮甘蓝型油菜。1991年,戚存扣通过对该组合杂种F1染色体减数分裂行为观察结果,认为将埃芥“CC”染色体组上的黄籽基因转移到甘蓝型油菜中是可能的。因为在杂种F1代,来自埃芥的和甘蓝型油菜的两组“CC”染色体组的染色体能完全配对。1987年,陈宝元提出用淡黄籽的甘蓝品种与含黄沙逊黄种皮基因的白菜型品种杂交,人工合成甘蓝型黄籽材料。黄籽油菜,尤其是甘蓝型黄籽油菜,将是今后油菜育种研究的主要课题之一。目前,有关油菜种皮色的形成机制还很不清楚,需要运用现代分析技术研究油菜种皮中的生理生化过程,以求发现与种皮色直接相关的化学物质或生理生化过程,为油菜种皮色的遗传控制提供基础。中国栽培的白菜型的、芥菜型的油菜中,存在大量的黄籽农家品种,对这些品种的收集、整理,将丰富黄籽油菜研究的内容。有关甘蓝型油菜种皮色遗传研究,还有许多问题有待解决,如为什么在甘蓝型油菜中不能获得纯黄种子?粒色遗传不稳定性的原因是什么?应用现代生物技术和遗传工程手段进行基因组代换,分子原位杂交,将外源种质的黄种皮基因转移到甘蓝型油菜中来,将是未来黄籽油菜育种研究的一个重要方面。【参考文献】:1 Vera C L,et al.Can Plant Scl.,1979,59:635~6372 Stringam G R.Can Plant Sci.,1980,60:331~3353 SchwetKa A.Theor Appl Genet,1982,62:161~1694 肖达人.作物学报,1983,8(4):245~2545 高永同.中国油料,1984,3:85~896 Shirzadegam M,G Robbelen.FETTE.SEIFEN,1985,6:235~2377 刘后利.作物杂志,1988,2:5~78 傅寿仲,唐继宏,戚存扣.江苏农业学报,1988,4(1):26~309 戚存扣,仲裕泉,等.江苏农业学报,1991,7(4):8~1210 王汉中.中国油料,1991,1:30~33(江苏省农业科学院戚存扣副研究员撰;傅寿仲审)