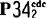 激酶
激酶【激酶】
细胞周期调节是一复杂过程。在酵母条件突变株中,已发现一类基因与细胞周期调节密切相关,称为细胞分裂周期基因(cdc基因)(L.H.Hartwell,1971),它们控制着细胞周期的启动及各时相的转换,其中以cdc2基因最重要。该基因是唯一在两个控制点G1→S、G2→M均起作用的cdc基因,它编码分子量为34kD的丝氨酸/苏氨酸蛋白激酶,P34cdc2激酶的活化是细胞分裂的增殖信号,它具有启动DNA复制和诱发有丝分裂的双重作用,在细胞周期调节中起重要作用。
20世纪70年代初,Masui等从蛙卵中发现一种能促进细胞成熟和分裂的因子,称为成熟促进因子(MPF)。1976年Nurse等发现和建立了cdc2酵母突变株,cdc2基因的编码产物即P34cdc2激酶,后确定两者为同一成分。P34cdc2激酶广泛存在于从酵母到人等多种生物中,形成一个庞大的家族。不同生物种族,其结构虽有差异,但具有很高的同源性。如酵母与人的有63%同源性(G.Draetta等,1987)。该酵母由催化亚基和调节亚基组成。催化亚基分子量为34kD,调节亚基又叫周期素。催化亚基由约300个氨基酸残基组成,含一般蛋白激酶的基本序列,同时又含该家族的特征序列,即:EGVPSTAFREISLLKE。催化亚基有3个重要的功能区(G.Draetta,1990):第1功能区在9~38位残基之间,是ATP的结合部位和该酶的活性部位;第2功能区在42~51位残基之间,是调节亚基的结合部位;第3+功能区在177~208位残基之间,是P13suc1的结合部位。P13suc1可与该酶高亲和力特异结合而使之广泛应用于实验研究,含P13suc的层析柱可抽提掉提取物中99%的P34cdc2激酶,且不影响其活性。此外,催化亚基的上14-苏氨酸、15-酪氨酸、33-赖氨酸、227-丝氨酸也具有非常重要的作用。调节亚基cyclin具有周期性积累和降解的特性,在细胞周期的特定时相与催化亚基结合。主要发现有3种cyclin:G1期cyclin、cyclin A、cyclin B。它们结构差异较大,但都含有一个相同的序列区域,即“cyclin box”。它们在细胞周期的不同时相发挥作用,G1期cyclin在G1末期与催化亚基结合,在G→S控制点起作用;cyclin A在S期和G2期与催化亚基结合,与DNA合成及S期向G2期的过渡有关;cyclin B在G2期末与催化亚基结合,可诱发有丝分裂。P34cdc2激酶对DNA复制和有丝分裂的诱发是必需的,在G1→S和G2→M两个控制点起中心调节作用。非洲爪蟾卵的提取物在体外能诱发DNA复制,但若将提取物中的P34cdc2激酶移走,则丧失诱发DNA复制的能力,说明P34cdc2激酶具有启动DNA复制的作用(J.J.Blow,1990)。但DNA复制的引发是一复杂过程,可能通过以下环节起作用:(1)对RNA聚合酶Ⅱ的调节作用:细胞由G1→S的移行需要RNA聚合酶Ⅱ指导下的基因转录,P34cdc2激酶可作用于RNA聚合酶Ⅱ,对其进行磷酸化修饰调节,P34cdc2激酶可通过促进RNA聚合酶Ⅱ指导下的基因转录,从而启动DNA复制(Pines等,1990)。(2)对SV40T抗原样蛋白的调节作用:把纯化的T抗原注入静止期细胞核中,可诱导DNA复制,T抗原带有使静止期细胞进入细胞周期的必要信息。R.Baserga认为,细胞内存在SV40T抗原样蛋白,并认为它是控制细胞增殖的主要成分。1989年Mcvey研究资料表明,T抗原如果不被磷酸化则无诱发DNA复制的作用。而体内外实验资料均证明T抗原是P34cdc2激酶的底物。P34cdc2激酶可能以细胞内T抗原样蛋白作为中间调节,以启动DNA复制。(3)对P53蛋白调节作用:P53是一个抗癌蛋白,有抑制细胞生长的作用,但突变的P53则具有促转化作用。P34cdc2激酶可磷酸化野生型P53(C.J.Marshall,1991),磷酸化的P53与SV40T抗原蛋白的结合能力降低,使之抑制复制的能力减弱。此外,还可使突变型的P53磷酸化,使之稳定性增加,有利于其发挥促增殖转化作用。(4)对高迁移率非组蛋白的DNA结合功能的影响:高迁移率非组蛋白(HMG)是一类丰富的核蛋白,可影响色体功能和DNA复制。HMG-1是其中的一个亚类,为DNA结合蛋白,结合到DNA的A-T丰富区,在增殖细胞中HMG-1的表达增加,它与细胞分裂和保持细胞的未分化状态密切相关。体内外实验资料均证明,HMG-1是P34cdc2激酶的有效底物,磷酸化部位是53位苏氨酸残基,该部位正是DNA结合的部位,它被磷酸化后则与DNA的结合能力下降。虽然HMG-1与DNA结合后所发挥的作用不清楚,但P34cdc2激酶对DNA结合蛋白的亲和力的影响可能是其启动DNA复制的机制之一(R.Reves,1991)。如c-myc、c-myb的编码产物是DNA结合蛋白,它们同样受到P34cdc2激酶的磷酸化修饰调节。(5)对核蛋白的调节作用:核仁蛋白是一种分子量为92kD的核蛋白,在核蛋白体RNA的合成和成熟过程起作用,它可直接作用于染色体,也可通过RNA聚合酶Ⅰ调节rRNA基因的转录。实验资料证明,核蛋白亦是P34cdc2激酶的底物,它的磷酸化可促进其功能作用,使rRNA的转录增加。此外,另一种核蛋白NO38也受到P34cdc2激酶的调节,NO38在核蛋白体的聚合和转运中起作用,它的磷酸化可影响它与核蛋白体RNA及蛋白质之间的相互作用。DNA的复制需一系列蛋白质因子的作用,P34cdc2激酶还可通过促进蛋白质的合成而促进DNA的合成(L.M.Boy等,1990)。有丝分裂是细胞一分为二的过程,其中包括一系列重要过程如染色质凝集,纺锤体形成、核膜破裂等。利用无细胞体系可直接观察到P34cdc2激酶对上述过程的促进,说明其具有诱发有丝分裂的作用。很早就观察到组蛋白H1以细胞周期依赖方式进行磷酸化,在G1晚期、G-端的一个丝氨酸残基被磷酸化;在S期,相邻的另外两个丝氨酸被磷酸化;进入M期后,C-端的一个丝氨酸残基又被磷酸化,组蛋白H1的磷酸化与染色质凝集密切相关。组蛋白H1是P34cdc激酶最早确定的底物。目前体外测定P34cdc2激酶活性实际上就是测定其组蛋白H1激酶活性(E.M.Bradbury等,1974)。在有丝分裂初期纺锤体形成,它的形成对于核模的破裂和染色体向两极的移动必不可少。纺鍾体由中心粒和微管组成,微管在中心粒周围呈放射状排列。P34cdc2激酶在G2期与中心粒结合,并催化中心粒蛋白磷酸化(E.Bailly等,1989),中心粒蛋白的磷酸化使微管的成核作用增强,而放射状排列在中心粒周围。P34cdc2激酶还可直接作用于微管(F.Verde等,1990),促进其在间隙的解聚及在分裂期的重组,还可维持其长度的稳定性,这可促进微管参与形成纺锤体,以及帮助染色体向两极的移动。核膜由内外两层组成,由3种蛋白质组成的核板与核内膜、染色质和核孔相连。核板在维持核的稳定性中起重要作用,此外还与染色质重组、DNA复制有关。板蛋白是丝状蛋白质,结构与中丝蛋白相似。在分裂开始时,核蛋白发生磷酸化,其溶解性增强促进核板解聚乃至核膜破裂(B.Luscher,1991)。P34cdc2激酶可催化板蛋白的磷酸化,促进核膜破裂。而在分裂中晚期,在特异磷酸酶的作用下,板蛋白脱磷酸,核板重新合成。波形纤维蛋白和索蛋白是细胞骨架的组成成份,前者存在于胞质中,形成从质膜到核膜的网架结构;后者在核内聚积于核膜下面。它们的解聚与核膜破裂密切相关,同样,它们的解聚也需磷酸化修饰调节。免疫沉淀法显示,从细胞中提取的波形纤维蛋白激酶就是P34cdc2激酶(T.C.Ned,1990),索蛋白也是该酶的底物。在细胞增殖分裂过程中,有一种蛋白发生高度磷酸化,即核蛋白体S6蛋白。胰岛素刺激细胞后使它的磷酸化程度急骤增强,S6蛋白的磷酸化可促进蛋白质合成。催化S6蛋白磷酸化的酶是S6激酶,该酶还可作用于核板蛋白,使之磷酸化而促进核膜破裂。它受PP60c-src磷酸修饰调节,而后者又是P34cdc2激酶的底物(D.Morgan等,1989),即P34cdc2激酶可间接促进S6激酶活性。P34cdc2-cyclin复合物有3种形式,它们的活性调节方式有差异,但基本方式相同,以P34cdc2-cyclinB为例介绍其活性调节。磷酸化/脱磷酸化修饰调节:与许多蛋白激酶相反,P34cdc2激酶磷酸化后失活,脱磷酸则活化。在间隙期,P34cdc2激酶14-苏氨酸和15-酪氨酸残基磷酸化,该酶处于失活状态。在进入M期前一瞬间,两部位脱磷酸,激酶迅速被活化且活性达峰值。在M中后期,调节亚基的解聚引起激酶失活(J.Gautier,1990)。目前还未分离出特异的P34cdc2激酶激酶和磷酸酶。据1991年K.Lundgren等研究资料表明,Wee和MIK1两基因的编码产物可能是P34cdc2激酶酪氨酸蛋白激酶,cdc25基因产物可激活P34cdc2激酶,它可能是磷酸酶的活化剂。调节亚基的调节作用:调节亚基的作用是多方面的。第一,它是该酶活化所必需的,在间隙期cyclin B不断合成积聚,并与磷酸化的催化亚基结合(R.Booher,1989),它的结合虽不直接使激酶活化,但是活化的必需前提,未与cyclin B结合的催化亚基即使脱磷酸也不能被活化;cylin B可能调节活性中心形成。第二,它调节激酶的底物特异性。Cyclin可能改变催化亚基的构象,使之与不同的底物相结合;也可能在底物与酶之间起桥梁作用,但只是推测,尚无实据。第三,cyclin的降解与激酶M期失活有关。进入M期后,由于特异的cyclin B磷酸酶和水解酶的出现,使cyclin B降解,调节亚基的解聚和降解使激酶失活。如果它的降解受阻,则可使激酶持续cdc2维持高活性。原癌基因c-mosxe的编码产物使cyclin B磷酸化,磷酸化后其稳定性增强使之不易被降解,因而c-mosxe过度表达则可引起P34cdc2激酶持续活化(H.Onkura等,1990),有丝分裂不能顺利完成,细胞停滞于M期。生长因子是细胞增殖所必需的,某些癌基因的活化可使细胞获得无限增殖的能力,它们与P34cdc2激酶的关系尚不很明了。生长因子与受体作用后,经信息传导,增殖信息转变为细胞内的化学信号,其作用的最后靶点是否就是P34cdc2激酶,通过它而诱发细胞增殖呢?1990年Barrett等报道,胰岛素刺激后,细胞内P34cdc2激酶活性升高,但这并不能说明两者之间有直接关系。许多癌基因的产物如PP60c-src、C-abl的产物等是P34cdc2激酶的底物,受到该酶的调节,于是人们推测,细胞分裂过程中细胞发生的一些变化如附着力减弱、细胞变圆等,可能是P34cdc2激酶作用于癌基因产物而引起的。此外,发现P21H-ras和c-mos1产物均可作用于P34cdc2激酶,使之活性升高,提示癌基因的活化可能启动细胞周期的调控机制,使细胞不断增殖分裂,这为解释癌基因在肿瘤发生中的作用提供了新的途径。SV40T抗原样蛋白有促进DNA复制起动的作用,有人认为抗癌基因产物P53与P105-Rb都可与其结合,从而抑制其促复制的作用。P34cdc2激酶可使P53和P105-Rb磷酸化,磷酸化后的P53和P105-Rb与SV40T抗原蛋白的结合能力降低,抑制复制的能力因之减弱。P34cdc2激酶的研究翻开了细胞周期调控研究新的一页,揭示了从酵母到人都存在相同的调节机制,但对其作用的环节还有待更深入的研究,如cdc2基因表达的调控、cdc2基因与其它cdc基因的关系,特别是生长因子、癌基因与P34cdc2激酶的关系更是广阔的研究领域。(北京医科大学基础医学院生物化学教研究室蔡家新、童坦君撰)