【红细胞免疫系统】
拼译:the red-cell iimmune system
红细胞有许多与免疫有关的物质(CR1、CR3、LFA-3、DAF、SOD酶等),数目众多,自成系统。红细胞有识别、粘附、浓缩、杀伤抗原、清除CIC的能力,参与机体免疫调控,并有完整的自我调控系统。许多疾病(如SLE)免疫发病机理中,红细胞免疫缺陷占有很重要的地位。该研究对全面认识机体免疫系统和免疫网络具有重要意义。
1930年杜克(L.H.Duke)发现锥虫在抗血清及补体存在时可粘附到人类红细胞上,并发现不同人的红细胞对锥虫的粘附能力高低不同。1953年纳尔逊(R.A.Nelson)发现人类红细胞与特异调理过的梅毒螺旋体及肺炎双球菌结合,称此为免疫粘附。推测红细胞膜存在免疫粘附受体,免疫复合物同该受体结合可促进白细胞的吞噬作用,并认为是宿主防御机制的一部分。1963年尼雪俄考(K.Nishioka)证实这种免疫粘附现象是通过人红细胞膜C3受体(现称第一补体受体,CR1)而实现的,1980年弗尔龙(D.T.Fearon)详细研究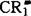 的性质,是分子量190000~250000的多态性膜糖蛋白。红细胞膜CR1密度为白细胞的
的性质,是分子量190000~250000的多态性膜糖蛋白。红细胞膜CR1密度为白细胞的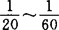 ,但血循环中红细胞数量却是白细胞的1000多倍,故CR总数85%以上在红细胞上。在同时期,加尔威(I.S.Garvey)发现新生兔红细胞有识别、储存牛血清白蛋白抗原的能力。1981年美国生殖免疫学家西格尔(I.Siegel)在前人研究的基础上发现红细胞有多种免疫功能,红细胞可粘附胸腺细胞,并发现血清中存在红细胞免疫粘附抑制因子,是一种不耐热的大分子的糖蛋白。在SLE患者中此因子增加,预见了血清中存在红细胞免疫调节系统。红细胞膜过氧化物酶活性与CR1活性有关,红细胞有杀伤致病原的效应细胞样作用。推测红细胞在阻止肿瘤细胞血行转移中有作用。用系统综合看待以往对红细胞免疫的研究成果,提出了“红细胞免疫系统”的新概念,冲破了传统上划分血细胞功能的“界限”,更新了人们对红细胞功能的认识。1982年梅多夫(M.1E.Medof)通过体外实验证明红细胞CR1和血浆中I因子共同作用将粘附的免疫复合物(IC)中的C3b降解为C3dg、C3d,而失去致炎性。1982年郭峰通过体外实验证明红细胞可粘附补体调理过的酵母菌,并证明SLE、肿瘤患者红细胞免疫粘附酵母菌的能力低下,该实验说明红细胞可直接粘附补体调理过的病原体。1983年科娜康夫(J.B.Cornacoff)在猴血循环内注入免疫复合物(IC),用同位素示踪,发现大多数IC很快与红细胞结合,并迅速被运至肝、脾,在该特定环境下,巨噬细胞膜上FC段受体比红细胞膜上CR1活性强,使IC从红细胞膜上脱落而被吞噬消毁。1986年郭峰通过体外对比实验证明红细胞可粘附补体调理过的各种肿瘤细胞,并证明肿瘤患者红细胞免疫粘附肿瘤细胞的能力下降,动物实验证明中药有效治疗后,红细胞免疫粘附肿瘤细胞能力增强。1988年威瑞拉(Virela)用单抗标记法证明红细胞膜有CR3。1990年派考德(J.P.Paccaud)采用折痕-标记免疫电镜比较PMN和红细胞的CR1形态,发现红细胞上几乎50%的CR1量≥3单位簇状分布,而这种簇状分布在PMN不到15%。CR1的这种簇状分布可使它与C3b包被的IC结合位点呈多价性,连接更为牢固。实验证明尽管单个白细胞表面CR1数(2500~6000/细胞)较红细胞(200~1000/细胞)多,但在细胞浓度相同时,两种细胞的IC结合率相同。而在CR1浓度等同的情况下,PNM与IC的结合量总是低于红细胞。由于血液中红细胞数远远超过白细胞,因而几乎所有补体调理过的抗原和IC都是由红细胞结合运至肝脾消毁的。红细胞免疫调控功能的研究报道有增多。1984年西格费索(A.Sigfuson)通过体外美洲商陆素刺激淋巴细胞转化实验发现加自身红细胞可增加淋巴细胞转化率和培养液中IgG、IgA量。1987年鲁杰利斯(M.T.Rugeles)发现自身红细胞加入人外周血单个核细胞培养管中可增强原发性和继发性特异抗体应答。1986年凯斯(L.Keyes)等发现人自身红细胞可增加T细胞产生γ-扰素。菲可液提取人外周血单个核细胞加适量PHA刺激诱导产生γ-干扰素。实验管加自身红细胞到淋巴细胞悬液中(10~50∶1)。发现加红细胞组γ-干扰素含量明显高于不加红细胞组,并且与加红细胞数有关,但与血型无关。用溶解的红细胞膜代替完整的红细胞仍观察到类似促进作用。抗CD2单抗可以抑制人红细胞和绵羊红细胞对T细胞产生γ-干扰素的促进作用。这种抗体可抑制淋巴细胞的增殖和红细胞与淋巴细胞之间的相互作用。用抗IL-2受体单抗研究发现红细胞可促进T细胞表达IL-2,增强T辅助细胞的免疫功能。1988年叶瑞拉(G.Yirella)通过体外抗淋巴细胞功能相关抗原3(LFA-3)单抗或抗CD2单抗处理和不处理的红细胞对促进B细胞增殖和免疫球蛋白的影响分析,认为红细胞的这种作用是同LFA-3与T细胞的CD2分子相互作用密切相关,推测是由于它促进T细胞IL-2受体表达和增强对外源性IL-2应答的敏感度所致,可能与增加B细胞生长因子或分化因子有关。表达LFA-3的红细胞有利于激活T辅助细胞。T辅助细胞除接受单一信号(加工后的抗原)刺激外,还接受第二信号——通过LFA3/CD2相互作用。1988年莎厄(H.Shau)在红细胞促进NK活性的对比研究中发现,当效靶细胞为80:1时,红细胞促NK细胞毒活性最高,红细胞与效应细胞比例以5:1时最强。1988年万内利(J.R.Yannelli)在培养瓶内加红细胞可促进LAK细胞的产量和活性,红细胞数与淋巴细胞数比为100:1时IL-2激活的LAK细胞活力最大。通过单抗阻断实验,证明这种促进作用与红细胞膜上LFA-3与淋巴细胞膜CD2相互作用密切相关。1985年福斯里德(J.Forslid)在体外通过对比实验证明红细胞促吞噬作用与红细胞CR1和SOD酶活性有关。1987年郭峰通过体外对比实验证明血清中还存在一种加热(58℃30min)不灭活的红细胞免疫粘附2进因子。1989年郭峰发现在体外对比实验中,红细胞和淋巴细胞或粒细胞可共同围攻粘附各种肿瘤细胞。从1930年发现锥虫在抗体及补体存在时粘附到人类红细胞上,到1981年提出“红细胞免疫系统”至今,研究发展迅速。回顾对淋巴细胞的认识,从认为是“终末细胞”到发现其有许多免疫功能,为现代免疫学奠定了基础。可以期望红细胞免疫系统的研究也将会有新的突破,将充实现代免疫学。红细胞免疫物质与遗传研究将会有新的进展,红细胞膜有β-内啡呔受体,神经内分泌与红细胞免疫关系已有初步报道;CR3与吞噬功能有关,红细胞膜上有CR3,是否说明红细胞对某些小抗原也有吞噬消毁作用;现已知CR1为2个等位基因共同编码,利用CR1基因探针对有核红细胞CR1基因缺陷进行诊断,在临床与基础理论研究方面将会有实用价值。红细胞免疫与白细胞免疫之间的关系研究,这是今后研究热点,红细胞可影响白细胞免疫功能;也有初步报道白细胞分泌的细胞因子(如IL-2干扰素)可促进红细胞免疫功能。但它们之间互相调控网络仍不清楚。红细胞免疫物质(如SOD酶)提纯应用于治疗、实现红细胞免疫功能测定方法多样化、实用化。将红免知识应用于临床诊治、中医药、以及其他生物学研究将有发展前景。【参考文献】:1 Siegel I,et al.The redcell immune system lancet,1981,2:5562 郭峰,等.红细胞免疫功能初步研究.中华医学杂志,1983,62(2):7153 Cornacoff J B,et al. Primate erythrocyte - immune complex -Clearing mechanism, J Clin Invest, 1983,71:2364 Medof ME. Complement - dependent maintenance of imma-ne Complex solability. In: Ratherk, Till Go, eds. The Complement system. New York Springer - Verlag, 1988. 4185 郭峰,等.天花粉对小鼠艾氏腹水癌的疗效和对红细胞免疫功能的影响,中西医结合杂志,1989,9(7):4186 Paccaud J P,et al. Difference in the clustering of complement receptor type I(CR1)on polymorphonaclear leukcytes and erythrocytes: effect on immune adherence Eur, J Immanol. ,1990,20:2837 郭峰,等.红细胞免疫及其调节功能测定方法,免疫学杂志,1990,6(1):608 Hebert LA, et al. Biologic significance of the erythrocyte Complement receptor- A primate perquisite. J lab Clin Med,1991,118(4):3019 郭峰,等.红细胞调控细胞免疫的事实与意义,上海免疫学杂志,1991,11(1):4910 郭峰.红细胞免疫系统研究展望,中国免疫学杂志,1991,7(增刊),2
,但血循环中红细胞数量却是白细胞的1000多倍,故CR总数85%以上在红细胞上。在同时期,加尔威(I.S.Garvey)发现新生兔红细胞有识别、储存牛血清白蛋白抗原的能力。1981年美国生殖免疫学家西格尔(I.Siegel)在前人研究的基础上发现红细胞有多种免疫功能,红细胞可粘附胸腺细胞,并发现血清中存在红细胞免疫粘附抑制因子,是一种不耐热的大分子的糖蛋白。在SLE患者中此因子增加,预见了血清中存在红细胞免疫调节系统。红细胞膜过氧化物酶活性与CR1活性有关,红细胞有杀伤致病原的效应细胞样作用。推测红细胞在阻止肿瘤细胞血行转移中有作用。用系统综合看待以往对红细胞免疫的研究成果,提出了“红细胞免疫系统”的新概念,冲破了传统上划分血细胞功能的“界限”,更新了人们对红细胞功能的认识。1982年梅多夫(M.1E.Medof)通过体外实验证明红细胞CR1和血浆中I因子共同作用将粘附的免疫复合物(IC)中的C3b降解为C3dg、C3d,而失去致炎性。1982年郭峰通过体外实验证明红细胞可粘附补体调理过的酵母菌,并证明SLE、肿瘤患者红细胞免疫粘附酵母菌的能力低下,该实验说明红细胞可直接粘附补体调理过的病原体。1983年科娜康夫(J.B.Cornacoff)在猴血循环内注入免疫复合物(IC),用同位素示踪,发现大多数IC很快与红细胞结合,并迅速被运至肝、脾,在该特定环境下,巨噬细胞膜上FC段受体比红细胞膜上CR1活性强,使IC从红细胞膜上脱落而被吞噬消毁。1986年郭峰通过体外对比实验证明红细胞可粘附补体调理过的各种肿瘤细胞,并证明肿瘤患者红细胞免疫粘附肿瘤细胞的能力下降,动物实验证明中药有效治疗后,红细胞免疫粘附肿瘤细胞能力增强。1988年威瑞拉(Virela)用单抗标记法证明红细胞膜有CR3。1990年派考德(J.P.Paccaud)采用折痕-标记免疫电镜比较PMN和红细胞的CR1形态,发现红细胞上几乎50%的CR1量≥3单位簇状分布,而这种簇状分布在PMN不到15%。CR1的这种簇状分布可使它与C3b包被的IC结合位点呈多价性,连接更为牢固。实验证明尽管单个白细胞表面CR1数(2500~6000/细胞)较红细胞(200~1000/细胞)多,但在细胞浓度相同时,两种细胞的IC结合率相同。而在CR1浓度等同的情况下,PNM与IC的结合量总是低于红细胞。由于血液中红细胞数远远超过白细胞,因而几乎所有补体调理过的抗原和IC都是由红细胞结合运至肝脾消毁的。红细胞免疫调控功能的研究报道有增多。1984年西格费索(A.Sigfuson)通过体外美洲商陆素刺激淋巴细胞转化实验发现加自身红细胞可增加淋巴细胞转化率和培养液中IgG、IgA量。1987年鲁杰利斯(M.T.Rugeles)发现自身红细胞加入人外周血单个核细胞培养管中可增强原发性和继发性特异抗体应答。1986年凯斯(L.Keyes)等发现人自身红细胞可增加T细胞产生γ-扰素。菲可液提取人外周血单个核细胞加适量PHA刺激诱导产生γ-干扰素。实验管加自身红细胞到淋巴细胞悬液中(10~50∶1)。发现加红细胞组γ-干扰素含量明显高于不加红细胞组,并且与加红细胞数有关,但与血型无关。用溶解的红细胞膜代替完整的红细胞仍观察到类似促进作用。抗CD2单抗可以抑制人红细胞和绵羊红细胞对T细胞产生γ-干扰素的促进作用。这种抗体可抑制淋巴细胞的增殖和红细胞与淋巴细胞之间的相互作用。用抗IL-2受体单抗研究发现红细胞可促进T细胞表达IL-2,增强T辅助细胞的免疫功能。1988年叶瑞拉(G.Yirella)通过体外抗淋巴细胞功能相关抗原3(LFA-3)单抗或抗CD2单抗处理和不处理的红细胞对促进B细胞增殖和免疫球蛋白的影响分析,认为红细胞的这种作用是同LFA-3与T细胞的CD2分子相互作用密切相关,推测是由于它促进T细胞IL-2受体表达和增强对外源性IL-2应答的敏感度所致,可能与增加B细胞生长因子或分化因子有关。表达LFA-3的红细胞有利于激活T辅助细胞。T辅助细胞除接受单一信号(加工后的抗原)刺激外,还接受第二信号——通过LFA3/CD2相互作用。1988年莎厄(H.Shau)在红细胞促进NK活性的对比研究中发现,当效靶细胞为80:1时,红细胞促NK细胞毒活性最高,红细胞与效应细胞比例以5:1时最强。1988年万内利(J.R.Yannelli)在培养瓶内加红细胞可促进LAK细胞的产量和活性,红细胞数与淋巴细胞数比为100:1时IL-2激活的LAK细胞活力最大。通过单抗阻断实验,证明这种促进作用与红细胞膜上LFA-3与淋巴细胞膜CD2相互作用密切相关。1985年福斯里德(J.Forslid)在体外通过对比实验证明红细胞促吞噬作用与红细胞CR1和SOD酶活性有关。1987年郭峰通过体外对比实验证明血清中还存在一种加热(58℃30min)不灭活的红细胞免疫粘附2进因子。1989年郭峰发现在体外对比实验中,红细胞和淋巴细胞或粒细胞可共同围攻粘附各种肿瘤细胞。从1930年发现锥虫在抗体及补体存在时粘附到人类红细胞上,到1981年提出“红细胞免疫系统”至今,研究发展迅速。回顾对淋巴细胞的认识,从认为是“终末细胞”到发现其有许多免疫功能,为现代免疫学奠定了基础。可以期望红细胞免疫系统的研究也将会有新的突破,将充实现代免疫学。红细胞免疫物质与遗传研究将会有新的进展,红细胞膜有β-内啡呔受体,神经内分泌与红细胞免疫关系已有初步报道;CR3与吞噬功能有关,红细胞膜上有CR3,是否说明红细胞对某些小抗原也有吞噬消毁作用;现已知CR1为2个等位基因共同编码,利用CR1基因探针对有核红细胞CR1基因缺陷进行诊断,在临床与基础理论研究方面将会有实用价值。红细胞免疫与白细胞免疫之间的关系研究,这是今后研究热点,红细胞可影响白细胞免疫功能;也有初步报道白细胞分泌的细胞因子(如IL-2干扰素)可促进红细胞免疫功能。但它们之间互相调控网络仍不清楚。红细胞免疫物质(如SOD酶)提纯应用于治疗、实现红细胞免疫功能测定方法多样化、实用化。将红免知识应用于临床诊治、中医药、以及其他生物学研究将有发展前景。【参考文献】:1 Siegel I,et al.The redcell immune system lancet,1981,2:5562 郭峰,等.红细胞免疫功能初步研究.中华医学杂志,1983,62(2):7153 Cornacoff J B,et al. Primate erythrocyte - immune complex -Clearing mechanism, J Clin Invest, 1983,71:2364 Medof ME. Complement - dependent maintenance of imma-ne Complex solability. In: Ratherk, Till Go, eds. The Complement system. New York Springer - Verlag, 1988. 4185 郭峰,等.天花粉对小鼠艾氏腹水癌的疗效和对红细胞免疫功能的影响,中西医结合杂志,1989,9(7):4186 Paccaud J P,et al. Difference in the clustering of complement receptor type I(CR1)on polymorphonaclear leukcytes and erythrocytes: effect on immune adherence Eur, J Immanol. ,1990,20:2837 郭峰,等.红细胞免疫及其调节功能测定方法,免疫学杂志,1990,6(1):608 Hebert LA, et al. Biologic significance of the erythrocyte Complement receptor- A primate perquisite. J lab Clin Med,1991,118(4):3019 郭峰,等.红细胞调控细胞免疫的事实与意义,上海免疫学杂志,1991,11(1):4910 郭峰.红细胞免疫系统研究展望,中国免疫学杂志,1991,7(增刊),2(第二军医大学附属长海医院免疫室郭峰研究员撰)