【肌肉收缩机制】
拼译:mechanism of muscle contraction
包括骨骼肌、心肌和平滑肌的肌肉组织,分别是躯体运动和一些重要内脏器官活动的动力来源。因此,阐明肌肉的收缩机制,对理解和运用躯体及某些器官的活动规律,是十分重要的。肌肉区分为上述3类,是根据其结构和收缩特性,但它们收缩产生力的机制,尤其是从分子水平看,是非常相似的。
自1930年起,相继提出肌肉收缩的乳酸说、磷酸肌酸说,以及后来的ATP初浅理论。至1939年,则发现肌肉收缩中肌蛋白的作用,但直到二次世界大战结束时,才将肌蛋白分离出,并认为肌纤蛋白与肌凝蛋白结合使ATP分解,并释放能量。当时的光学显微镜不能分辨其细微结构,直到20世纪50年代初期,应用电子显微镜观察到有两类相互重叠的肌丝,一类是肌凝蛋白组成的粗肌丝,一类是肌纤蛋白组成的细肌丝。后来电镜进一步显示,肌肉收缩时,两类肌丝均不缩短,因此产生了肌丝滑行理论。肌纤蛋白聚合成双螺旋状,成为细肌丝的主干。多个肌凝蛋白的尾部,组成粗肌丝的主干,其头部突出于粗肌丝的表面,称为横桥,它有与肌纤蛋白结合的部位,还有与ATP结合的部位。滑行是横桥与肌纤蛋白分子结合并摆动,牵拉细肌丝沿粗肌丝长轴方向滑动(故称横桥理论),滑行产生的张力与两类肌丝重叠的程度成正比。肌纤蛋白与肌凝蛋白结合后,使后者被激活,作为ATP酶而分解ATP。当ATP分解产物被清除,横桥与新的ATP结合,使横桥与肌纤蛋白的连接断开,并与下一个肌纤蛋白结合,周而复始进行连接和脱开的循环,而产生滑动,使肌肉缩短。1957年赫克斯利(A.F.H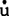 xley)对上述横桥理论的建立做出突出贡献,其后在20世纪60年代和70年代,尽管许多人提出各自的解释,但均未改变上述基本理论。进一步研究指出,肌凝蛋白包括大分子量的重酶解肌凝蛋白(HMM)和小分子量的轻酶解肌凝蛋白(LMM),前者又可酶解为2类碎片,HMM-S-1和HMM-S-2。HMM-S-1组成横桥的双头结构,HMM-S-2和LMM组成肌凝蛋白的尾部。肌纤蛋白中,G肌纤蛋白为球形分子,F肌纤蛋白是包含G肌纤蛋白的纤维状结构,它组成了肌纤蛋白的双螺旋结构。上述肌肉收缩机制中,化学能转变为机械能,是分子水平研究中的最热烈的课题。这种能量的转变就是指ATP的作用模式,在1971年莱蒙(R.W.Iymn)和泰勒(E.W.Taylor)提出的ATP作用模式假设的基础上,1986年希波(M.G.Hibberd)等进一步补充与完善这个作用模式,指出肌凝蛋白分子上的ATP分解,实际是在肌凝蛋白横桥与肌纤蛋白连接之前,ATP分解产生的ADP和无机磷(pi),仍束缚于肌凝蛋白上。而ATP分解释放的化学能,被转移至肌凝蛋白(M)上,产生高能形式的肌凝蛋白
xley)对上述横桥理论的建立做出突出贡献,其后在20世纪60年代和70年代,尽管许多人提出各自的解释,但均未改变上述基本理论。进一步研究指出,肌凝蛋白包括大分子量的重酶解肌凝蛋白(HMM)和小分子量的轻酶解肌凝蛋白(LMM),前者又可酶解为2类碎片,HMM-S-1和HMM-S-2。HMM-S-1组成横桥的双头结构,HMM-S-2和LMM组成肌凝蛋白的尾部。肌纤蛋白中,G肌纤蛋白为球形分子,F肌纤蛋白是包含G肌纤蛋白的纤维状结构,它组成了肌纤蛋白的双螺旋结构。上述肌肉收缩机制中,化学能转变为机械能,是分子水平研究中的最热烈的课题。这种能量的转变就是指ATP的作用模式,在1971年莱蒙(R.W.Iymn)和泰勒(E.W.Taylor)提出的ATP作用模式假设的基础上,1986年希波(M.G.Hibberd)等进一步补充与完善这个作用模式,指出肌凝蛋白分子上的ATP分解,实际是在肌凝蛋白横桥与肌纤蛋白连接之前,ATP分解产生的ADP和无机磷(pi),仍束缚于肌凝蛋白上。而ATP分解释放的化学能,被转移至肌凝蛋白(M)上,产生高能形式的肌凝蛋白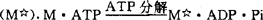 高能形式的肌凝蛋白继而与肌纤蛋白(A)结合,触发贮在肌凝蛋白横桥中的能量释放,产生横桥摆动,同时ADP与Pi也从肌凝蛋白释放出来。
高能形式的肌凝蛋白继而与肌纤蛋白(A)结合,触发贮在肌凝蛋白横桥中的能量释放,产生横桥摆动,同时ADP与Pi也从肌凝蛋白释放出来。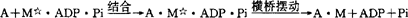 在肌肉收缩时,每个横桥要发生多次循环活动,即反复连接和脱开,每一循环伴随一分子ATP的分解。肌凝蛋白横桥与肌纤蛋白的连接很牢固,必须由一个新的ATP分子结合至肌凝蛋白,才能使横桥脱开。
在肌肉收缩时,每个横桥要发生多次循环活动,即反复连接和脱开,每一循环伴随一分子ATP的分解。肌凝蛋白横桥与肌纤蛋白的连接很牢固,必须由一个新的ATP分子结合至肌凝蛋白,才能使横桥脱开。 如此反复循环。肌肉收缩的分子机制的研究过程,曾先是离体状态在溶液中的研究,分析了粗细肌丝的蛋白组成,以及ATP水解过程中,各种成分相互反应的步骤。从分子水平进行这样的研究,自然是非常重要的,但这只是从生物化学角度的阐述,探讨在肌肉纤维中的反应过程与在溶液中的结果的一致性,是多年来的研究方向。戈德曼(Y.E.Goldman)等1984年首先应用Caged ATP技术和18O交换技术,分析肌肉纤维生化反应,证明肌肉纤维内的反应特征,在许多方面相同于溶液中研究的结果。布伦纳(B.Brenner)应用高精度测量机械作用的技术,来研究横桥动力学变化,并应用X线衍射技术,来观察肌肉纤维横桥在不同功能状态下的结构变化。结果表明,在肌肉纤维松驰时,横桥具有类似于在溶液中称为弱结合状态下所现示的特征,即肌纤蛋白与横桥亲和力弱;这种结合不依赖于Ca2+的存在;二者连接和脱开迅速达到平衡。不同的是,在溶液中连接和脱开的速度是相同的,而在肌肉纤维中有一定变化范围。在肌肉松驰时仍然存在少量的横桥与肌纤蛋白连接的事实,似乎与通常称肌肉收缩产生与否,是由于横桥连接的发生和阻断的理论有矛盾。当然对肌肉收缩的进一步的控制机制,目前尚有不同看法。在肌肉纤维活动时,横桥处于另一种状态,它具有类似于在溶液中称为强结合状态下所显示的特征,即肌纤蛋白与横桥亲和力强;Ca2+加速二者的结合;二者的连接和脱开达到平衡,而脱开比较慢。不同的是,在溶液中连接和脱开的速度相同,而在肌肉纤维中,脱开的速度也有一定变化范围。横桥的形态在弱结合时与强结合时也是不同的,这是因为横桥的构型不同,从弱结合到强结合构型的变化,是摆动和产生力量的关键。为了在生理或接近生理状态下,观察肌肉纤维的横桥方位和摆动方式,1987年托马斯(D.D.Thomas)采用光谱探测技术。应用这种探测技术,可直接测出横桥在肌肉收缩时产生摆动,而且横桥的摆动有一定的变化范围,证实了肌丝滑行理论。自然,这些新技术所显示的结果,还存在难以解释的疑问。以往也有作者应用双折射方法,根据肌丝重叠程度影响折射强度的原理,探讨分析完整肌肉组织在收缩时,肌凝蛋白分子的方位。还曾有作者用荧光物质标记肌凝蛋白,镜下观察肌凝蛋白分子结构。从20世纪50年代提出肌肉收缩机制的肌丝滑行理论后,许多研究工作者进一步探讨了肌丝滑行的详细机制。在此过程中,他们应用现代科学技术,采用许多新的研究方法,力图从机械收缩、生化反应、形态结构甚至能量产生探讨其机制,取得很大进展。今后,从生理学、生化学、形态学和分子遗传学方面研究肌肉收缩机制,仍为其研究方向,尤其是分子遗传学的发展,并应用于肌肉收缩研究中,会给这个题目的研究带来冲击。随着许多边缘学科的发展,许多内容和方法必然渗透到肌肉收缩的研究当中,与许多学科的发展交织在一起,今后随着相关学科的发展,将会给肌肉收缩的研究带来新的活力。
如此反复循环。肌肉收缩的分子机制的研究过程,曾先是离体状态在溶液中的研究,分析了粗细肌丝的蛋白组成,以及ATP水解过程中,各种成分相互反应的步骤。从分子水平进行这样的研究,自然是非常重要的,但这只是从生物化学角度的阐述,探讨在肌肉纤维中的反应过程与在溶液中的结果的一致性,是多年来的研究方向。戈德曼(Y.E.Goldman)等1984年首先应用Caged ATP技术和18O交换技术,分析肌肉纤维生化反应,证明肌肉纤维内的反应特征,在许多方面相同于溶液中研究的结果。布伦纳(B.Brenner)应用高精度测量机械作用的技术,来研究横桥动力学变化,并应用X线衍射技术,来观察肌肉纤维横桥在不同功能状态下的结构变化。结果表明,在肌肉纤维松驰时,横桥具有类似于在溶液中称为弱结合状态下所现示的特征,即肌纤蛋白与横桥亲和力弱;这种结合不依赖于Ca2+的存在;二者连接和脱开迅速达到平衡。不同的是,在溶液中连接和脱开的速度是相同的,而在肌肉纤维中有一定变化范围。在肌肉松驰时仍然存在少量的横桥与肌纤蛋白连接的事实,似乎与通常称肌肉收缩产生与否,是由于横桥连接的发生和阻断的理论有矛盾。当然对肌肉收缩的进一步的控制机制,目前尚有不同看法。在肌肉纤维活动时,横桥处于另一种状态,它具有类似于在溶液中称为强结合状态下所显示的特征,即肌纤蛋白与横桥亲和力强;Ca2+加速二者的结合;二者的连接和脱开达到平衡,而脱开比较慢。不同的是,在溶液中连接和脱开的速度相同,而在肌肉纤维中,脱开的速度也有一定变化范围。横桥的形态在弱结合时与强结合时也是不同的,这是因为横桥的构型不同,从弱结合到强结合构型的变化,是摆动和产生力量的关键。为了在生理或接近生理状态下,观察肌肉纤维的横桥方位和摆动方式,1987年托马斯(D.D.Thomas)采用光谱探测技术。应用这种探测技术,可直接测出横桥在肌肉收缩时产生摆动,而且横桥的摆动有一定的变化范围,证实了肌丝滑行理论。自然,这些新技术所显示的结果,还存在难以解释的疑问。以往也有作者应用双折射方法,根据肌丝重叠程度影响折射强度的原理,探讨分析完整肌肉组织在收缩时,肌凝蛋白分子的方位。还曾有作者用荧光物质标记肌凝蛋白,镜下观察肌凝蛋白分子结构。从20世纪50年代提出肌肉收缩机制的肌丝滑行理论后,许多研究工作者进一步探讨了肌丝滑行的详细机制。在此过程中,他们应用现代科学技术,采用许多新的研究方法,力图从机械收缩、生化反应、形态结构甚至能量产生探讨其机制,取得很大进展。今后,从生理学、生化学、形态学和分子遗传学方面研究肌肉收缩机制,仍为其研究方向,尤其是分子遗传学的发展,并应用于肌肉收缩研究中,会给这个题目的研究带来冲击。随着许多边缘学科的发展,许多内容和方法必然渗透到肌肉收缩的研究当中,与许多学科的发展交织在一起,今后随着相关学科的发展,将会给肌肉收缩的研究带来新的活力。(山东医科大学刘京璋教授撰)