【植物的硝酸还原酶】
拼译:nitrate reductase in plants
植物吸收利用的无机氮有NH4+和NO3-(主要是NO3-)。NO3-进入植物体内的同化过程是:外界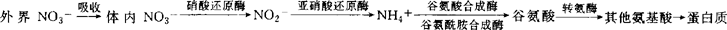 。硝酸还原酶(简称NR)是这一过程的第一限速酶,它具有以下特点:(1)植物中显著的诱导酶之一,易被诱变得到突变体;(2)酶结构复杂,由多个亚基组成,含有不同组分酶,其活性易受体内外因素影响;(3)是氮素代谢的关键酶,影响农作物的总氮和蛋白氮水平,与作物的耐肥性有密切关系,并同时具有功能多样性的特点,对植物的其它代谢过程如能量代谢、铁离子同化与运转、水分胁迫、光呼吸、氯离子的还原、分子氧(O2)的分解释放等也有重要影响。NR在植物基因表达、蛋白质分子基础研究和植物代谢途径及调控研究中占有十分重要的地位。
。硝酸还原酶(简称NR)是这一过程的第一限速酶,它具有以下特点:(1)植物中显著的诱导酶之一,易被诱变得到突变体;(2)酶结构复杂,由多个亚基组成,含有不同组分酶,其活性易受体内外因素影响;(3)是氮素代谢的关键酶,影响农作物的总氮和蛋白氮水平,与作物的耐肥性有密切关系,并同时具有功能多样性的特点,对植物的其它代谢过程如能量代谢、铁离子同化与运转、水分胁迫、光呼吸、氯离子的还原、分子氧(O2)的分解释放等也有重要影响。NR在植物基因表达、蛋白质分子基础研究和植物代谢途径及调控研究中占有十分重要的地位。
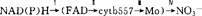 。除作为电子载体外,部位Ⅰ还具有脱氢酶的功能,能利用NAD(P)H还原NO3-、Fe[CN]3、哺乳类细胞色素C等,NR对Fe3+的还原功能可能对植物体内铁离子代谢起着重要作用,该部位能被巯基试剂(ρ-HMB)强烈抑制;部位Ⅳ(末端NR)能利用FADH2或还原态MV还原NO3-,氰化物和叠氮化物是该部位的底物竞争性抑制剂。Solomonson(1986)获得该酶的两个多肽片段,Ⅰ:分子量较小,含FAD,能利用NADH还原Fe(CN)3;Ⅱ:分子量较大,含Mo元素,可利用还原态MV还原NO3-,这直接证明NR有两个活性中心。NR催化反应的双位“乒乓”稳态动力学机制也支持该假说。不过,在某些植物中还发现其他动力学机制,如随机序列快速平衡机制、底物随机结合和产物“乒乓”释放机制等。就NR的氨基酸组成而言,通过对烟草、大麦、玉米等基因的克隆及其核苷酸顺序的确定,再由上述几种NR基因推译出的氨基酸顺序有80%的同源性。此外,将烟草NR基因推译出的氨基酸顺序与已知顺序的其它蛋白质比较,发现NR的cytb557和脱氢酶组分(包括活性中心Ⅰ)分别与哺乳类细胞内质网上的cytb5和NADH-cytb5还原酶(一种黄素脱氢酶)惊人的相似。这些结果将有助于NR生化特性的深入研究。NR是植物的诱导酶,对NR的诱导机理有酶蛋白从头合成和酶前体活化两种观点。Funkhouser等(1983)认为小球藻中存在NR前体,并已分离得到酶的脱辅基蛋白——酶前体。张德颐也报告了有酶前体的存在,在非诱导条件下植物组织含有一定的NR活力也是NR前体存在的佐证。Somers等(1983)用火箭免疫电泳技术证明
。除作为电子载体外,部位Ⅰ还具有脱氢酶的功能,能利用NAD(P)H还原NO3-、Fe[CN]3、哺乳类细胞色素C等,NR对Fe3+的还原功能可能对植物体内铁离子代谢起着重要作用,该部位能被巯基试剂(ρ-HMB)强烈抑制;部位Ⅳ(末端NR)能利用FADH2或还原态MV还原NO3-,氰化物和叠氮化物是该部位的底物竞争性抑制剂。Solomonson(1986)获得该酶的两个多肽片段,Ⅰ:分子量较小,含FAD,能利用NADH还原Fe(CN)3;Ⅱ:分子量较大,含Mo元素,可利用还原态MV还原NO3-,这直接证明NR有两个活性中心。NR催化反应的双位“乒乓”稳态动力学机制也支持该假说。不过,在某些植物中还发现其他动力学机制,如随机序列快速平衡机制、底物随机结合和产物“乒乓”释放机制等。就NR的氨基酸组成而言,通过对烟草、大麦、玉米等基因的克隆及其核苷酸顺序的确定,再由上述几种NR基因推译出的氨基酸顺序有80%的同源性。此外,将烟草NR基因推译出的氨基酸顺序与已知顺序的其它蛋白质比较,发现NR的cytb557和脱氢酶组分(包括活性中心Ⅰ)分别与哺乳类细胞内质网上的cytb5和NADH-cytb5还原酶(一种黄素脱氢酶)惊人的相似。这些结果将有助于NR生化特性的深入研究。NR是植物的诱导酶,对NR的诱导机理有酶蛋白从头合成和酶前体活化两种观点。Funkhouser等(1983)认为小球藻中存在NR前体,并已分离得到酶的脱辅基蛋白——酶前体。张德颐也报告了有酶前体的存在,在非诱导条件下植物组织含有一定的NR活力也是NR前体存在的佐证。Somers等(1983)用火箭免疫电泳技术证明 诱导NR活性从零逐渐增大,与NR蛋白质水平增高呈平行关系,否定了酶前体的存在。Campbell(1986)进一步用酶联免疫吸附法(ELISA)发现玉米NR蛋白质合成先于NR活性的出现。由此看来,NO3-诱导NR蛋白的合成,酶活的表达受其他因子影响。进一步深入到核酸水平研究发现NO3-的主要作用在于NR基因表达的启动,光则可能主要通过光敏素系统作用于NR-mRNA的转录。有人推测,NO3-可能通过与一种NR调节蛋白结合,然后插入NR基因附近的核苷酸序列,活化NR基因表达的启动子,使NR-mRNA被有效地转录。也有人认为NO3-可能不是直接活化NR基因的效应物,而硝酸盐同化过程中的某些中间产物可能与NR基因活化有关。20世纪70年代以来,用细胞突变体的方法,己从烟草、大麦等植物中得到两类NR-突变体,钼辅因子基因突变型和脱辅基蛋白基因突变型,它们的杂种细胞又可恢复NR活力,表明它们的基因型是互补的。Miller(1983)的烟草NR-突变体杂交试验结果指出,NR受到两个不连锁基因的控制,它们就是酶的脱辅基蛋白结构基因。Sherrard(1976)等指出,小麦中7B和7D两条染色体参与NR活力的调节。林振武(1989)报道,水稻NR诱导形成受到基因转录的控制,认为至少有两个基因位点参与酶活力的调节,一个是酶的编码基因,参与酶的合成;另一个是酶钝化蛋白基因,参与酶的衰减。NR蛋白的合成调节可发生在基因水平、转录水平和翻译水平上;NR翻译后的活性调节则主要通过酶的活化与钝化、氧化与还原两个系统来调节。现已从高等植物中分离得到NR稳定因子、活化因子、钝化因子和抑制因子,它们对NR都具有特异性。引起酶失活的钝化蛋白有NR结合蛋白和NR水解蛋白,对NR专一性的水解蛋白已分别从玉米和小麦中分离出来,植物体内NR有氧化(活性态)和还原两种状态,体内许多代谢物(如NADH)影响该酶两种状态的可逆转变,从而调节NR活性。NR极易受体内外环境因素的影响。NO3-的吸收与输送以及从贮存库中的释放,直接影响NO3-的底物效应;光、pH值、植物激素和有机酸对NR活性也有很大影响。低βH作用于NR的从头合成,而不是对酶前体的简单活化,作用机理尚不清楚,至于植物激素,一般认为6BA促进NR活性而GA、ABA则抑制NR活性;但也有ABA促进NR活性的报道,植物激素在调节NR活性中的作用值得深入研究。NR的研究正由细胞水平不断深入到分子水平,成为被充分研究的植物酶之一。可预料以下几方面将是研究的热点:(1)NR结构与特性的深入研究,包括NR的初级结构、高级结构及其催化机制,植物种间NR差异及其在进化上的意义;(2)NR基因及其表达,利用分子克隆技术.开展NR基因分离、鉴定和克隆、表达的研究;(3)NR诱导机制和活性调节,利用分子生物学技术,在NR基因水平、转录水平和翻译水平上进行研究;(4)NR功能多样性的进一步研究:(5)NR的应用研究,如NR与生化育种、NR与作物固氮以及最终采用遗传工程手段利用NR基因改变NR活力,调节植物的氮代谢,从而提高作物的产量与品质。【参考文献】:1 Miguel G,et al. Annual Review of Plant Physiology. 1981, 32:1692 Sinha S K. The Physiology and Biochemistry of Drought R esistance in Plants. 1981,145-1693 Kleinhofs A, Warner R L,Narayanan K R. Molecular,Cell B,ology 1985,2:91-1214 汤玉玮,林振武,陈敬祥.中国农业科学,1985,6:39~455 林振武.植物生理学通讯,1987,6:8~136 Wilbur H.Physiologia Plantarum 1988,74:214~2197 林振武,汤玉玮.中国科学(B辑),1989,4:379~3858 Srivastava H S.Phytochemistry.1992,31:2974
诱导NR活性从零逐渐增大,与NR蛋白质水平增高呈平行关系,否定了酶前体的存在。Campbell(1986)进一步用酶联免疫吸附法(ELISA)发现玉米NR蛋白质合成先于NR活性的出现。由此看来,NO3-诱导NR蛋白的合成,酶活的表达受其他因子影响。进一步深入到核酸水平研究发现NO3-的主要作用在于NR基因表达的启动,光则可能主要通过光敏素系统作用于NR-mRNA的转录。有人推测,NO3-可能通过与一种NR调节蛋白结合,然后插入NR基因附近的核苷酸序列,活化NR基因表达的启动子,使NR-mRNA被有效地转录。也有人认为NO3-可能不是直接活化NR基因的效应物,而硝酸盐同化过程中的某些中间产物可能与NR基因活化有关。20世纪70年代以来,用细胞突变体的方法,己从烟草、大麦等植物中得到两类NR-突变体,钼辅因子基因突变型和脱辅基蛋白基因突变型,它们的杂种细胞又可恢复NR活力,表明它们的基因型是互补的。Miller(1983)的烟草NR-突变体杂交试验结果指出,NR受到两个不连锁基因的控制,它们就是酶的脱辅基蛋白结构基因。Sherrard(1976)等指出,小麦中7B和7D两条染色体参与NR活力的调节。林振武(1989)报道,水稻NR诱导形成受到基因转录的控制,认为至少有两个基因位点参与酶活力的调节,一个是酶的编码基因,参与酶的合成;另一个是酶钝化蛋白基因,参与酶的衰减。NR蛋白的合成调节可发生在基因水平、转录水平和翻译水平上;NR翻译后的活性调节则主要通过酶的活化与钝化、氧化与还原两个系统来调节。现已从高等植物中分离得到NR稳定因子、活化因子、钝化因子和抑制因子,它们对NR都具有特异性。引起酶失活的钝化蛋白有NR结合蛋白和NR水解蛋白,对NR专一性的水解蛋白已分别从玉米和小麦中分离出来,植物体内NR有氧化(活性态)和还原两种状态,体内许多代谢物(如NADH)影响该酶两种状态的可逆转变,从而调节NR活性。NR极易受体内外环境因素的影响。NO3-的吸收与输送以及从贮存库中的释放,直接影响NO3-的底物效应;光、pH值、植物激素和有机酸对NR活性也有很大影响。低βH作用于NR的从头合成,而不是对酶前体的简单活化,作用机理尚不清楚,至于植物激素,一般认为6BA促进NR活性而GA、ABA则抑制NR活性;但也有ABA促进NR活性的报道,植物激素在调节NR活性中的作用值得深入研究。NR的研究正由细胞水平不断深入到分子水平,成为被充分研究的植物酶之一。可预料以下几方面将是研究的热点:(1)NR结构与特性的深入研究,包括NR的初级结构、高级结构及其催化机制,植物种间NR差异及其在进化上的意义;(2)NR基因及其表达,利用分子克隆技术.开展NR基因分离、鉴定和克隆、表达的研究;(3)NR诱导机制和活性调节,利用分子生物学技术,在NR基因水平、转录水平和翻译水平上进行研究;(4)NR功能多样性的进一步研究:(5)NR的应用研究,如NR与生化育种、NR与作物固氮以及最终采用遗传工程手段利用NR基因改变NR活力,调节植物的氮代谢,从而提高作物的产量与品质。【参考文献】:1 Miguel G,et al. Annual Review of Plant Physiology. 1981, 32:1692 Sinha S K. The Physiology and Biochemistry of Drought R esistance in Plants. 1981,145-1693 Kleinhofs A, Warner R L,Narayanan K R. Molecular,Cell B,ology 1985,2:91-1214 汤玉玮,林振武,陈敬祥.中国农业科学,1985,6:39~455 林振武.植物生理学通讯,1987,6:8~136 Wilbur H.Physiologia Plantarum 1988,74:214~2197 林振武,汤玉玮.中国科学(B辑),1989,4:379~3858 Srivastava H S.Phytochemistry.1992,31:2974(安徽师范大学周阮宝副教授撰)