【大豆同工酶类】
拼译:soybean isozymes
同工酶指分子结构及理化性质不同而催化作用相同的酶,它代表着基因的多型性。同工酶广泛存在于植物的不同种类、不同品种和不同器官以及生长发育的不同时期。研究同工酶对于探究大豆遗传育种和变异、起源和进化、分类以及生长发育和病理生理等方面有着理论意义和实践意义。在大豆中已发现有28种同工酶。
大豆同工酶电泳位点的遗传模式及定位研究对遗传育种工作中的基因连锁图谱的建立非常重要,它能帮助解决大豆图谱非常不完全的问题。1968年,巴特里(Buttery)和巴泽尔(Buzzell)首先用电泳法发现不同大豆品种种皮的过氧化物酶(POD)活性有高低之分,后来证实这是由一对基因控制的。显性基因(Ep)决定高活性,而隐性基因(ep)决定低活性。1978年,戈尔曼(M.B.Gorman)和基安格(Y.T.Kiang)发现乙醇脱氢酶(ADH)具有4~5个基因控制的遗传模式,淀粉酶及四唑氧化酶也具有各自的遗传控制模式。1983年,他们又研究另外8种同工酶。到1991年止,人们已先后对24种大豆同工酶进行了电泳位点的遗传模式的研究(基安格等,1987、1989;杜恩格J.Y.H.Doong,1988)。人们围绕大豆同工酶之间以及它们与其他抗病及形态学性状之间的连锁关系做了大量的测定工作。1980年,希尔德布兰德(D.F.Hildebrand)等人首先报道了Ap(酸性磷酸酶)与Ti(胰蛋白酶抑制剂)连锁,属于大豆连锁群9。之后,人们在研究中发现Ap与Lap1(亮氨酸氨肽酶)、Pgd1(磷酸葡糖酸脱氢酶)与Pgi1(磷酸葡萄糖异构酶)、Lap1与Ti、ACO3(乌头酸酶)与Sp1(淀粉酶的一个位点)连锁,还发现Ep与Fr1,Adh1与W1(花色)、Idh1(异柠檬酸脱氢酶)与F及Rj1(结瘤反应性状)、ACO2与Rj2等都有连锁关系。所有研究中其它各项均是独立遗传的。在品种分类及鉴定方面,开展工作最早的是巴特里和巴泽尔,他们根据大豆种皮中POD活性高低将大豆分成两大类。戈尔曼、基安格及奇安格等人则在这方面做了大量较为系统的工作。他们在1982年建立了大豆12个同工酶酶谱类型图,并利用此同工酶系统将253个供试品种中的168个鉴定出来。1987年,杜恩格与基安格等人建立了大豆16个同工酶谱类型图,列出367个品种的8种同工酶谱类型。因这些研究工作为大豆遗传育种及品种分类、鉴定提供了有用的依据而得到应用。1980年,韦特(Wetter)发现大豆-烟草体细胞杂种中天门冬氨酸氨基转移酶与ADH同工酶谱皆具有来自双亲的一些酶带。刘学军、苗以农(1991)则发现具有杂种优势的大豆后代辽豆3号及吉林21其POD同工酶谱具有偏父现象。基安格等人(1983)还认为由同工酶位点计算出的遗传距离可成为选择成功父母本的指标。刘德璞等人(1989)发现DNA导入后引起突变的材料的SOD(超氧物歧化酶)同工酶谱具有供体的一些同工酶谱带,证明DNA导入的成功及在突变育种上的可能性。人们还利用同工酶技术发现了分别缺失L-1、L-2、L-3以及同时缺失L1和L2,L2和L3(L:脂肪氧化酶)的大豆材料,也获得β-AM和脲酶分别缺失的品种(希尔德布兰德,1981;喜多村,1983、1986)。这些材料的发现对大豆遗传育种和上述酶类生理作用的研究很有意义。同工酶技术的应用在研究栽培大豆的起源、进化、分布及其与野生大豆的关系方面也取得一些进展。1982年,海莫威茨(T.Hymowitz)等人测定了SBTi-A2和Sp1电泳位点在15个亚洲国家和地区大豆群中的分布,结合电泳结果和其它方面的资料指出,来自亚洲的大豆可分为3个基因中心,包含7个种质库,认为大豆是由中国传向其它亚洲国家的。奇安格与戈尔曼(1981,1984)则发现野生大豆无论是杂合频率还是酶多态水平都高于栽培大豆,认为这是由于野生大豆要适应各种复杂环境,而栽培大豆的酶多态性则由于长期驯化而损失。布罗茨(S.L.Broich)、虞京葳(1983)及徐豹等分别对大豆Ep基因频率、酯酶酶谱及SOD活性的分析表明,野生菌种>中间型>栽培种;徐豹(1990)还提出大豆SOD变异的可能途径。大豆同工酶类生理作用研究较多的有脂肪氧化酶(LOX)、POD和SOD。大豆含有大量的LOX,它使大豆产生豆腥味,降低了大豆制品的食用价值和营养价值。这是由于LOX引起酶促脂质过氧化作用所致。有人认为LOX还与衰老有关,其活性随植株生长而增加,并在加速衰老实验中随种子活力下降而降低。但旺(J.YWang),1990年的研究资料表明,LOX的3个同工酶带之一,L-2可能在保持种子活力及大豆生长方面起一定作用,而不是促进衰老,从而认为LOX并不是种子衰老的原因。A.Kaeperska 1985年报道由LOX催化的多不饱和脂肪酸的氧化的加强,导致ACC产生的乙烯的增加。但LOX确切的生理功能还不十分清楚。POD同工酶是人们研究最多而功能却非常不明确的一种酶。它广泛存在于植物体内,可能参与调节吲哚乙酸水平、乙烯的形成、细胞壁形成,调节膜的透性、脂肪酸氧化,控制O2透过种皮的种子休眠及抗病等多种生理过程,有人认为它可作为细胞分化的指标。在对大豆POD同工酶的研究中,齐哥德勒(J.A.Zygadlo)1982年,刘学军、苗以农于1990、1991年发现大豆各器官在生育过程中表现具有特殊差异的特有同工酶谱。衰老和人工衰老引起不同POD同工酶谱带的消失或迁移率发生变化,种子活力也下降(A.S.Rao,1986)。种子中POD主要分布在种皮中,可能与种皮的硬化、抵抗传染病及种子的贮存质量有关(巴特里等,1968;戈尔曼,1976)。对POD与抗病的关系研究较多。拉马拉基(N.V.Rameraje,Urs)1974年报道感病的大豆植株同健康植株相比,POD活性要增强,抗病品种要高于不抗病品种。宋英淑(1986)则发现抗旱性越强的品种,其叶片POD活性的相对变化越小。SOD是清除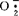 专一性的酶。近几年来,它在高等植物的氧伤害、器官衰老及抗逆性中所起的防护作用已逐渐获得证实。大豆,随种子成熟,其SOD活性增强,但各谱带在种子发育过程中同步表达(庄炳昌,1986)。1990年,罗广华等发现高氧促使大豆幼苗内产生
专一性的酶。近几年来,它在高等植物的氧伤害、器官衰老及抗逆性中所起的防护作用已逐渐获得证实。大豆,随种子成熟,其SOD活性增强,但各谱带在种子发育过程中同步表达(庄炳昌,1986)。1990年,罗广华等发现高氧促使大豆幼苗内产生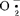 速率增快,同时又使幼苗内SOD活性水平提高以减轻
速率增快,同时又使幼苗内SOD活性水平提高以减轻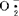 增高引起的细胞伤害。梁峥等人1991年发现,聚乙二醇处理种子后,SOD活性高于受低温吸胀冷害的种子子叶的活性,认为SOD保护正在修补的膜系统免遭伤害。同工酶的研究和利用显示着广阔的前景和强大的生命力,但还有许多工作要做。大豆的遗传连锁图谱非常不完全,还需在同工酶分离分析技术进一步完善的基础上做大量细致的工作。未来10~20年,多种同工酶系统将成为大豆品种的鉴定、分类及育种工作中非常重要的工具,使这些方面的工作向分子水平发展。各种同工酶尤其是同工酶谱中各条带的生理功能还远未搞清楚,但这方面的工作将继续进行并可能有所突破。【参考文献】:1 Buttery BR.R.I. Buzzell Crop Science. 1968,8:722~7252 Gorman M B. Jornal of Heredity. 1989 ,69(4) ;255~2583 Kiang Y T,Gorman M B. Soybean. In Isozymes in Plant Genetics and Breeding Part B. 1983,295~3284 Wetter L R. Chromosome and isozyme studies on cells derived from protoplast fusion of Nicotiana glauca with Glycine max - Nicotiana glauca cell, hybrids Theoretical and Applied Genetics. 1980,57(6) = 273-2765 梁峥,等.植物生理学报,1990,32(7):538~5436 徐豹,等.植物学报,1990,32(7):538~5437 刘学军,苗以农.中国油料,1992,1:12~14
增高引起的细胞伤害。梁峥等人1991年发现,聚乙二醇处理种子后,SOD活性高于受低温吸胀冷害的种子子叶的活性,认为SOD保护正在修补的膜系统免遭伤害。同工酶的研究和利用显示着广阔的前景和强大的生命力,但还有许多工作要做。大豆的遗传连锁图谱非常不完全,还需在同工酶分离分析技术进一步完善的基础上做大量细致的工作。未来10~20年,多种同工酶系统将成为大豆品种的鉴定、分类及育种工作中非常重要的工具,使这些方面的工作向分子水平发展。各种同工酶尤其是同工酶谱中各条带的生理功能还远未搞清楚,但这方面的工作将继续进行并可能有所突破。【参考文献】:1 Buttery BR.R.I. Buzzell Crop Science. 1968,8:722~7252 Gorman M B. Jornal of Heredity. 1989 ,69(4) ;255~2583 Kiang Y T,Gorman M B. Soybean. In Isozymes in Plant Genetics and Breeding Part B. 1983,295~3284 Wetter L R. Chromosome and isozyme studies on cells derived from protoplast fusion of Nicotiana glauca with Glycine max - Nicotiana glauca cell, hybrids Theoretical and Applied Genetics. 1980,57(6) = 273-2765 梁峥,等.植物生理学报,1990,32(7):538~5436 徐豹,等.植物学报,1990,32(7):538~5437 刘学军,苗以农.中国油料,1992,1:12~14(东北师范大学刘学军撰)