【水稻白叶枯病抗性遗传】
拼译:genetics of resistane to bacterial blight in rice
它是通过遗传研究发现不同的抗性基因和掌据病菌致病性同水稻抗性互作关系中的遗传变异特点,从而实现根据病原菌的消长合理替换抗性基因的科学的抗病育种之要求。因此,抗病性的遗传研究在促进抗病育种的发展和充实这一领域的基础理论方面起着十分重要的作用。 日本最先进行水稻白叶枯病抗性遗传研究。1961年,西村报道由一对显性基因控制黄玉等的抗性,该基因位于第11染色体。1966年,鹫尾(Washio)报道农林27和关东60的抗性受制于一对显性基因,早生爱国3号由一对显性基因控制日本当时所有的菌群。1968年,国际水稻研究所赫尤(Heu)等报道Sigadis携有一不完全显性抗性基因。1972年,墨蒂(Murty)和库什(Khush)报道BJI携有一对隐性抗性基因;DZ192抗性似乎由2对隐性基因制约。1967年,坂口报道Xa-1和Xa-2分别控制日本菌群Ⅰ和Ⅱ的抗性,都位于第11染色体,以2%~16%重组值连锁。江塚等1975年报道早生爱国3号群品种在成株期有一对显性基因控制日本菌群Ⅰ、Ⅱ和Ⅲ的抗性,命名为Xa-w,后改为Xa-3。国际水稻研究所1976~1979用菲律宾当时的1个流行菌系(属小种1)鉴定了6个抗性基因。1977年帕特皮西特(PatPisit)等报道了全生育期抗病的一对不完全显性基因Xa-4和一对隐性基因Xa-5,两者独立遗传。1976年利比洛约(Librojo)报道了Saemora Mangga有一对与Xa-4等位的基因,因为是成株期抗病,命名为Xa-4b。1978年西德赫(Sidhu)等报道Zenith和Malagkit Sungsong等品种携有另1个成株抗性基因Xa-6;1978、1979年Sidhu等又发现DV85和DZ78等除携有xa-5外尚有1个制约在花期抗性的显性基因xa-7,并报道P1231129携有抗性基因xa-8。1983年辛(Singh)等报道Sateng携有一对成株抗性基因xa-9.1983年吉村在国际水稻研究所开始用菲律宾当时的4个生理小种进行遗传研究,发现抗小种2的鉴别品种Cas209携有显性基因Xa-10。1986年小川和山本报道RP9-3等品种携有显性基因Xa-11控制日本小种Ⅱ和ⅢA的抗性。1987年小川等将黄玉和Java14抗印度尼西亚小种V的一显性基因Xa-Kg重新命名为Xa-12。除Xa-12外,上述抗病基因都采用本地菌系,不能直接比较。1982~1987年,国际水稻研究所与日本合作,统一用双方菌系比较了初期系统鉴定的抗性基因。1989年小川和Khush报道的结果表明,几个成株抗性基因Xa-4b,Xa-6和Xa-9与Xa-3是等位的,从而删除了早先命名的上述3个基因。1990年,Khush等和井田等发现1个来自长药野生稻的全生育期抗印度和菲律宾所有小种的抗源,携有一对完全显性抗性基因Xa-21。在此之前,其他8个新基因Xa-13>Xa-20都已报道。1987年小川等报道BJ1群有两对隐性基因Xa-5和新基因Xa-13。迄今已报道了21个国际命名的白叶枯病抗性基因,所用菌系、抗性范围等概括如表:表1 系统命名的稻白叶枯病抗性基因 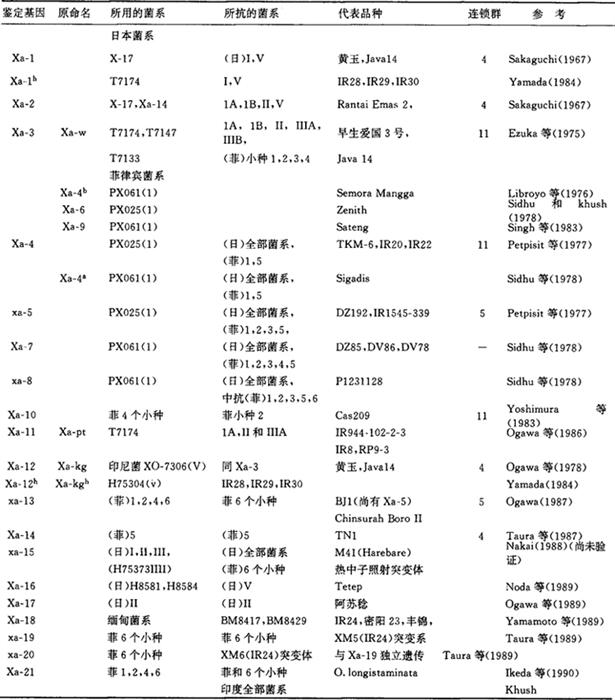
1977年,中山大学生物系观察33个杂交组合F1的抗性表现为显性或不完全显性;1978年,章琦分析73个杂交组合F1的抗性为显性、不完全显性和隐性现象,认为子1代的抗性取决于供体的基因型。这一领域的研究随着抗病育种工作的开展而深入。1989年,谢岳峰等报道15个云南地方品种对菌系江陵691的抗性由1~2对基因制约,若干个基因于当时测定的已知基因非等位。戴陆国等报道,毫梅携有新基因Xa-i(t),与第Ⅳ+Ⅸ连锁群即第2染色体的n1-1以重组值37.1%连锁。余功新等报道,对菲小种1携有Xa-4基因的IR28对我国菌系江陵691和OS75的抗性分别由Xa-a和Xa-h制约,它们属Ⅷ连锁群。1991年,万建民、朱立宏研究8个太湖晚梗品种对4个中国菌系的抗性遗传,认为品种与菌系间有明显的特异性互作;抗病性受感病亲本遗传背景的影响。许多研究证实同一个品种在不同菌系作用下的遗传行为各不相同。这些研究报道反映了寄主与病原菌的生物间遗传学的特性。1986~1991年,章琦等与国际水稻研究所合作采用国际基因命名的菌系分析了我国的主要抗源的基因组成和系谱渊源,揭示了我国抗性基因利用单一的问题。籼稻主要是Xa-4,梗稻集中于Xa-3[5,6]。伍尚忠等1988年报道了华南籼稻品种华竹矮等均有Xa-4的亲缘。抗源遗传变异狭窄,挖掘和利用新基因已是当务之急。籼粳亚种间杂交后代的抗性遗传表现较为复杂。1985年,万建民、朱立宏报道DV85的抗性在不同亚种间杂交后代的遗传表有特异性。1988年,周毓珍、朱立宏观察DV85的衍生中间抗源在不同遗传背景下的抗性转育效应不同。1988年,张耀钢、朱立宏报道7个籼稻中间抗源在不同亚种遗传背景下的抗性遗传行为无明显差别。育种后代的抗性转育效应也是抗病育种家需要深入了解的问题。建立我国水稻白叶枯病抗性遗传系统研究体系和基因命名的标准化,为新抗源的挖掘和利用提供一套完善的遗传学分析体系,是深化这一领域研究工作的基础。21个国际抗性基因中,除Xa-21外都来自栽培稻,仅少数具有育种价值,开发和利用野生稻中丰富的抗白叶枯病新基因,将有很大的发展前景。细胞遗传结合生物技术基因工程,探讨克服野栽杂交不育和基因导入的途径将是这一领域较为活跃的前沿课题。RFLP技术在稻病抗性基因定位上的应用已是国际研究热点。如果以常规育种为基础,将分子遗传技术应用于基因图谱和抗病育种,将使抗病遗传育种的理论和应用出现新的突破,产生深远的意义。【参考文献】:1 Nishmura y,Bull Natl lnst.Agric.Sci.,1961,D9:171~2352 Washio O,Kariya k,Toriyama k,Bull.chugoku Agric.Exp.sth.,1966,A13:55~853 Sakaguchi S.Bull Natl lust Agric Sci.,1967,D16:1~184 Petpisit V,khush G S,kauffman H E.Crop lmprov.,1979,7(17):551~5545 Sidhu G S,khush GS,Mew TW.Crop lmprov.,1979,6:19~256 Yoshimura A,Mew T W,et al.phytopathology.,1983,73:1409~14127 Ogawa T,Khush GS.Bacterial Blight of Rice.International Rice Research lnstitule,Manila,philippines,1989,177~1928 谢岳峰,等.作物遗传研究通讯,1989,5~89 Khush G S,et al.Rice Genetic Newsletter,1990,121~12210 章琦.中国农业科学,1991,24(3):26~36(中国农业科学院作物所章琦研究员撰) |