【鸟类的原生殖细胞】
拼译:primordial germ cells in the aves
鸟类原生殖细胞是由胚盘的上胚层细胞形成的,它以较大的体积和大量的糖元极易与体细胞区别开。如同其他脊椎动物一样,鸟的原生殖细胞内也有类似生殖质的生殖颗粒。鸟类原生殖细胞具有运动能力,基迁移方向与生殖腺原基吸引能力有关,与胚体内细胞外基质的合成在时空上有密切的关系。它是一种研究转向决定和转基因研究的一个较为理想的实验材料。
鸟类原生殖细胞,当它在组织或未分化性腺中常有伪足(T.Fujimot,1976),因此在不同时期其形状略有不同。由于原生殖细胞有大量糖元,所以用PAS方法即可识别。细胞质内有大量细胞器,在原生殖细胞进入运动之前细胞质内有大量的微管和成束的微丝,它们主要分布于质膜附近,这就奠定了它具有运动能力的结构基础(刘荣秀,1990)。在胚胎发育第10期,位于生殖新月区的原生殖细胞常呈灶状集聚,相邻的原生殖细胞有桥粒样结构,可能与同步成熟有关(刘荣秀,1990)。在细胞核内缘有少量异染色质,常染色质则为均匀分布。有1~2个核仁,光镜观察核为实线条状嗜派洛宁染色。如以RNA酶处理,核仁则不着色。根据嗜派洛宁染色和RNA酶处理后不着色的结果,表明核仁成分为核糖核酸,但它能否表示是真正的核仁尚待进一步研究(T,Fujimoto,1976)。1987年张天荫报道,在鸟类原生殖细胞中还没有找到生殖质一类的物质。刘荣秀(1990)报道,孵育36小时位于生殖新月区的原生殖细胞,其细胞核内、核膜间及胞质有一种圆形的电子致密颗粒(电子较密小体)。这些颗粒由点状电子致密点构成,埋于特别细的盘曲成海绵状纤维之中,其间充满基质,结构和其他动物的生殖颗粒极为相似,它是由核内产生,进入核周池,借移膜破裂方式进入细胞质的。这与无尾类动物的情况十分相似(S.Holwill等,1987)。P.D.Nieuwkoop等(1979)指出,鸟类原生殖细胞来自胚胎早期的初级下胚层。根据自80年代以来的研究资料证明,它不是来自下胚层而是来自上胚层。H.Eyal-Giladi等(1981)将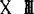 期鸡和鹌鹑胚盘上下胚层作嵌合胚胎,体外培养,当孵育到7~8期,用Feulgen染核异染色质作标记,证明原生殖细胞来自上胚层。(L.A.Sutasurya 1983)等通过分别培养
期鸡和鹌鹑胚盘上下胚层作嵌合胚胎,体外培养,当孵育到7~8期,用Feulgen染核异染色质作标记,证明原生殖细胞来自上胚层。(L.A.Sutasurya 1983)等通过分别培养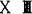 期-4期鸡胚上下胚层证明,在原条形成时原生殖细胞自上胚层逐步移向下胚层,在4期时还有许多原生殖细胞仍位于上胚层中,到6期时其数目则相当少。L.E.Urven等(1988)利用一种抗小鼠胚胎肿瘤细胞单抗EMA-1作标记,通过免疫组化方法证明鸡胚原生殖细胞来自上胚层。这种标记方法时间短,并更可靠,一直可保持到配子发生前(W.Loveless等1990);以一种低聚糖抗原FC10·2作标记也证明鸡胚原生殖细胞是来自上胚层的。M.Ginsberg等(1987)将Ⅹ-Ⅻ期胚盘透明区的中央盘、边缘带和暗区进行分割培养,结果表明鸡胚原生殖细胞主要来自胚盘透明区的中央盘,并证明早在Ⅹ期就已决定。当然,不同的鸟类孵育前原生殖细胞在透明区分布的位置略有不同,鸭分布不规律,鹌鹑则位于胚盘的前半部(D.Cuminge等,1989)。当原生殖细胞从上胚层移到下胚层后,由于下胚层的形态发生运动将它带到生殖新月区内。在4~8期原生殖细胞从生殖新月区的内胚层脱离开,位于内外胚层之间。当10期胚外中胚层侵入生殖新月区并开始形成血岛时,它就进入血管。此时胚胎前端已形成血管内,在12期可于胚外血管中找到原生殖细胞,13期胚胎开始心跳和血液循环(D.P.Nieuwkoop等,1979)。15期原生殖细胞在卵黄动脉后1.2mm处进入相当加厚的脏壁中层体腔上皮,并入原生殖细胞的加厚上皮向将来生殖腺方向移动,16期后通过体腔角向体壁中层方向移动,最后到18期以加厚上皮位于体腔角和中肾之间即将来生殖嵴处。这表示开始并入原生殖细胞加厚的脏壁中层上皮成为生殖嵴表面的上皮(A.Nakamura等,1987)。约在孵育3d形成未分化性腺。M.Nakamura(1988)统计,在正常发育情况下约有20%原生殖细胞不进入生殖腺而到达别的组织中.其中90%是进入头部神经管四周的间叶组织,这也许与头部微血管网发育早也较发达以及血流较慢,导致“误入歧途”的。为了探索原生殖细胞迁移的原因,Simon将带有生殖新月区胚盘前半部侧连于带有生殖腺原基胚胎的后半部,结果前者的原生殖细胞通过血液循环进入后者的生殖腺原基;如将去除生殖新月区的胚盘同正常胚盘作连体,结果两个胚胎的生殖腺中均有原生殖细胞,这表明原生殖细胞是通过血液循环从胚外进入胚内生殖腺的(P.D.Nieuwkoop等,1979),并认为可能是生殖腺上皮产生一种物质吸引原生殖细胞。为了证实后一种情况,T.Fuimoto等(1984)将两天到两天半胚胎的原生殖细胞和同期生殖嵴原基一起培养,发现原生殖细胞向生殖嵴迁移,最终到达生殖嵴原基。Kawana等(1986)进一步将原生殖细胞与同期几种原基(神经管、心脏、尿囊、肝和生殖嵴)作等距离共培养,证明原生殖细胞向生殖嵴原基作定向移动,表明生殖嵴确具一种吸引原生殖细胞的能力。如将原生殖细胞培养于缺乏生殖嵴原基的胶原蛋白培养基上,它就不运动,如带有生殖嵴原基就能产生伪足(T.Kawana等,1987)。这种生殖腺原基具有长距离吸引原生殖细胞的能力。在哺乳类动物也得到证明;还发现在培养基中有生殖嵴可使体外培养的原生殖细胞在培养物中消失的时间延长。(I.Godin等,1990)Rogufska等(1971)将小鼠后肠移到鸡胚生殖腺区,发现鸡胚生殖腺上皮也可以吸引小鼠的原生殖细胞,表明这种吸引能力还没有物种的区别。至于生殖腺原基分泌什么物质吸引原生殖细胞的迁移,W.J.Swartz(1975)认为是一种类固醇物质,其根据是他将一种雄激素或雌激素注入孵育33h的鸡胚,由于激素的干扰,所在检查孵育到5d的鸡胚生殖孵时发现其中原生殖细胞数大为减少。这种意见还没有得到更多学者的证实。原生殖细胞本身具有变形运动的能力。如将14~16期的原生殖细胞培养于有鸡胚背系膜的培养基上,其运动速度平均为26μm/h,最大可达58μm/h,其迁移方向由下面的间叶细胞长轴的方向决定(T.Kawana等,1984)。这与无尾两栖类动物和哺乳类动物的情况十分相似。Y.Ando等(1983)在体内找到原生殖细胞正通过微血管内皮间隙进入胚内脏壁中层之间的图像,这些原生殖细胞先粘附在血管内表面,然后产生一个膨大的伪足状突起通过血管壁内皮的缝隙进入脏壁中层。M.Nakamura等(1990)将10期胚胎后部1/3切去,当孵育到18期,集中于胚体的原生殖细胞有90%位于头部神经管四周;他们认为由于缺乏生殖腺,头部血管内皮和间叶细胞可能对在血液循环中的原生殖细胞有一种亲和能力,导致它们集中于头部。所有羊膜动物的原生殖细胞最终一定要集中并通过肠背系膜而到达生殖腺原基,而它在胚体组织间的迁移与细胞外基质密切有关。E.Urven等(1989)将抗片层蛋白、纤连蛋白、硫酸软骨素、蛋白多糖和胶原蛋白Ⅳ标记细胞外基质证明,原生殖细胞的迁移与基质在时空上的出现密切有关。这种时空关系在无尾两栖类动物和哺乳类动物均已得到证实。无论由生殖质决定还是由诱导而形成的原生殖细胞,它们都是在通过一定的迁移途径到达生殖嵴的过程中实现其分化的。因此,许多学者通过异位移植的原生殖细胞来研究决定了的细胞是否一定按其原定方向分化为生殖细胞。M.Nakamura等(1988,1991)虽在鸡胚正常发育的或切去生殖腺的原基中都发现有许多原生殖细胞进入胚胎头部神经管四周的间叶细胞,由于他们没有进一步研究其后来的命运,所以还不了解其分化情况。但是,人们在无尾两栖类动物已取得许多结果。C.Wylie等(1985)将爪蟾45期胚胎的的原生殖细胞标记后注入囊胚后期(9期)的囊胚腔中,结果它可分化为3个胚层的细胞,表明原生殖细胞的分化是由其最终所处的部位决定的,也表明它是一种多潜能细胞。移核实验资料也证明原生殖细胞核的多潜能性比同期其它体细胞大得多。L.D.Smith,(1965)J.H.Cleine(1986)将爪蟾受精卵倒置后发现所发育的胚胎许多原生殖细胞位于前肠,这些异位的原生殖细胞则分化为食道的或肠的上皮细胞。K.Ikenishi等(1988)将3H-胸腺嘧啶标记的原生殖细胞移植到神经胚内胚层的前半部和后半部,结果前者的原生殖细胞由于无法迁移到生殖嵴,以后就分化为食道的、胃的和肠的上皮,只有后者可迁移到生殖嵴而分化为生殖细胞。因此,他们认为对于原生殖细胞的分化,内胚层特定的部位是一个必不可少的条件。这种受到不同于原来外界环境的影响而向不同于原来的方向进行分化的转向决定,原生殖细胞可能是一种很好的研究对象。B.C.Wentworth等(1989)提出将鸟类原生殖细胞作为一种有用的基因转移工具。他用采用紫外线照射早期鸡胚的生殖新月区域或以二甲磺酸丁酯注入鸡胚杀死原生殖细胞以及用雄鸡和雌鹌鹑交配的方法获得不育的胚胎,然后将所需研究的原生殖细胞注入不育胚胎来研究其发育和子代的情况。他们将显性的野生鹌鹑的原生殖细胞作为施主注入不育的隐性白羽鹌鹑胚胎受体中,得到两个白色雌性的和一个白色雄性的子代,然后分别再与隐性白羽鹌鹑交配,结果在其子代中获得白羽的和体披野生型羽毛的杂合性子代。J.N.Petitte等(1990)将芦花洛克鸡胚Ⅹ期胚盘注入白来抗鸡胚胎的胚下腔,53个实验中获得6个羽毛表现型为嵌合体胚胎,其中一个孵化为雄性,他们把它与芦花洛克母鸡交配,其目的为测定施主细胞并入宿主睾丸中达到何种程度,在获得719个后代中有两个表现型为芦花洛克,表明施主细胞中的原生殖细胞已并入宿主胚胎的睾丸中,从其血液和精子DNA指纹图像来看不同于来抗鸡。从嵌合体血液DNA指纹图像来看仅为芦花洛克所有。因此,实验结果表明分离芦花洛克X期胚盘细胞提供了来自神经嵴的色素细胞、红血球和生殖细胞。所以,他们认为这种移植胚盘细胞技术可能有助于禽类转基因以及通过体外培养建立多潜能干细胞的研究。【参考文献】:1 Eyal-Giladi H,et al.J Embryo Exp Morph.1981,65:1392 张天荫.细胞生物学杂志,1987,4:1453 张天荫.原生殖细胞(丁汉波等主编·发育生物学).北京:高等教育出版社,19874 Urven E,et al.Anat Rec.,1989,224:245 Wentworth B C,et al.Poultry Sci.1989,68:7996 Loveless W,et al.Development,1990,108:977 刘荣秀.动物学报,1990,36:1148 Petitle J N,et al.Development,1990,108:1859 Godin I,et al.Developnent,1990,108:35710 Nakamura m,et al.Anat Rec,1991,229:109
期-4期鸡胚上下胚层证明,在原条形成时原生殖细胞自上胚层逐步移向下胚层,在4期时还有许多原生殖细胞仍位于上胚层中,到6期时其数目则相当少。L.E.Urven等(1988)利用一种抗小鼠胚胎肿瘤细胞单抗EMA-1作标记,通过免疫组化方法证明鸡胚原生殖细胞来自上胚层。这种标记方法时间短,并更可靠,一直可保持到配子发生前(W.Loveless等1990);以一种低聚糖抗原FC10·2作标记也证明鸡胚原生殖细胞是来自上胚层的。M.Ginsberg等(1987)将Ⅹ-Ⅻ期胚盘透明区的中央盘、边缘带和暗区进行分割培养,结果表明鸡胚原生殖细胞主要来自胚盘透明区的中央盘,并证明早在Ⅹ期就已决定。当然,不同的鸟类孵育前原生殖细胞在透明区分布的位置略有不同,鸭分布不规律,鹌鹑则位于胚盘的前半部(D.Cuminge等,1989)。当原生殖细胞从上胚层移到下胚层后,由于下胚层的形态发生运动将它带到生殖新月区内。在4~8期原生殖细胞从生殖新月区的内胚层脱离开,位于内外胚层之间。当10期胚外中胚层侵入生殖新月区并开始形成血岛时,它就进入血管。此时胚胎前端已形成血管内,在12期可于胚外血管中找到原生殖细胞,13期胚胎开始心跳和血液循环(D.P.Nieuwkoop等,1979)。15期原生殖细胞在卵黄动脉后1.2mm处进入相当加厚的脏壁中层体腔上皮,并入原生殖细胞的加厚上皮向将来生殖腺方向移动,16期后通过体腔角向体壁中层方向移动,最后到18期以加厚上皮位于体腔角和中肾之间即将来生殖嵴处。这表示开始并入原生殖细胞加厚的脏壁中层上皮成为生殖嵴表面的上皮(A.Nakamura等,1987)。约在孵育3d形成未分化性腺。M.Nakamura(1988)统计,在正常发育情况下约有20%原生殖细胞不进入生殖腺而到达别的组织中.其中90%是进入头部神经管四周的间叶组织,这也许与头部微血管网发育早也较发达以及血流较慢,导致“误入歧途”的。为了探索原生殖细胞迁移的原因,Simon将带有生殖新月区胚盘前半部侧连于带有生殖腺原基胚胎的后半部,结果前者的原生殖细胞通过血液循环进入后者的生殖腺原基;如将去除生殖新月区的胚盘同正常胚盘作连体,结果两个胚胎的生殖腺中均有原生殖细胞,这表明原生殖细胞是通过血液循环从胚外进入胚内生殖腺的(P.D.Nieuwkoop等,1979),并认为可能是生殖腺上皮产生一种物质吸引原生殖细胞。为了证实后一种情况,T.Fuimoto等(1984)将两天到两天半胚胎的原生殖细胞和同期生殖嵴原基一起培养,发现原生殖细胞向生殖嵴迁移,最终到达生殖嵴原基。Kawana等(1986)进一步将原生殖细胞与同期几种原基(神经管、心脏、尿囊、肝和生殖嵴)作等距离共培养,证明原生殖细胞向生殖嵴原基作定向移动,表明生殖嵴确具一种吸引原生殖细胞的能力。如将原生殖细胞培养于缺乏生殖嵴原基的胶原蛋白培养基上,它就不运动,如带有生殖嵴原基就能产生伪足(T.Kawana等,1987)。这种生殖腺原基具有长距离吸引原生殖细胞的能力。在哺乳类动物也得到证明;还发现在培养基中有生殖嵴可使体外培养的原生殖细胞在培养物中消失的时间延长。(I.Godin等,1990)Rogufska等(1971)将小鼠后肠移到鸡胚生殖腺区,发现鸡胚生殖腺上皮也可以吸引小鼠的原生殖细胞,表明这种吸引能力还没有物种的区别。至于生殖腺原基分泌什么物质吸引原生殖细胞的迁移,W.J.Swartz(1975)认为是一种类固醇物质,其根据是他将一种雄激素或雌激素注入孵育33h的鸡胚,由于激素的干扰,所在检查孵育到5d的鸡胚生殖孵时发现其中原生殖细胞数大为减少。这种意见还没有得到更多学者的证实。原生殖细胞本身具有变形运动的能力。如将14~16期的原生殖细胞培养于有鸡胚背系膜的培养基上,其运动速度平均为26μm/h,最大可达58μm/h,其迁移方向由下面的间叶细胞长轴的方向决定(T.Kawana等,1984)。这与无尾两栖类动物和哺乳类动物的情况十分相似。Y.Ando等(1983)在体内找到原生殖细胞正通过微血管内皮间隙进入胚内脏壁中层之间的图像,这些原生殖细胞先粘附在血管内表面,然后产生一个膨大的伪足状突起通过血管壁内皮的缝隙进入脏壁中层。M.Nakamura等(1990)将10期胚胎后部1/3切去,当孵育到18期,集中于胚体的原生殖细胞有90%位于头部神经管四周;他们认为由于缺乏生殖腺,头部血管内皮和间叶细胞可能对在血液循环中的原生殖细胞有一种亲和能力,导致它们集中于头部。所有羊膜动物的原生殖细胞最终一定要集中并通过肠背系膜而到达生殖腺原基,而它在胚体组织间的迁移与细胞外基质密切有关。E.Urven等(1989)将抗片层蛋白、纤连蛋白、硫酸软骨素、蛋白多糖和胶原蛋白Ⅳ标记细胞外基质证明,原生殖细胞的迁移与基质在时空上的出现密切有关。这种时空关系在无尾两栖类动物和哺乳类动物均已得到证实。无论由生殖质决定还是由诱导而形成的原生殖细胞,它们都是在通过一定的迁移途径到达生殖嵴的过程中实现其分化的。因此,许多学者通过异位移植的原生殖细胞来研究决定了的细胞是否一定按其原定方向分化为生殖细胞。M.Nakamura等(1988,1991)虽在鸡胚正常发育的或切去生殖腺的原基中都发现有许多原生殖细胞进入胚胎头部神经管四周的间叶细胞,由于他们没有进一步研究其后来的命运,所以还不了解其分化情况。但是,人们在无尾两栖类动物已取得许多结果。C.Wylie等(1985)将爪蟾45期胚胎的的原生殖细胞标记后注入囊胚后期(9期)的囊胚腔中,结果它可分化为3个胚层的细胞,表明原生殖细胞的分化是由其最终所处的部位决定的,也表明它是一种多潜能细胞。移核实验资料也证明原生殖细胞核的多潜能性比同期其它体细胞大得多。L.D.Smith,(1965)J.H.Cleine(1986)将爪蟾受精卵倒置后发现所发育的胚胎许多原生殖细胞位于前肠,这些异位的原生殖细胞则分化为食道的或肠的上皮细胞。K.Ikenishi等(1988)将3H-胸腺嘧啶标记的原生殖细胞移植到神经胚内胚层的前半部和后半部,结果前者的原生殖细胞由于无法迁移到生殖嵴,以后就分化为食道的、胃的和肠的上皮,只有后者可迁移到生殖嵴而分化为生殖细胞。因此,他们认为对于原生殖细胞的分化,内胚层特定的部位是一个必不可少的条件。这种受到不同于原来外界环境的影响而向不同于原来的方向进行分化的转向决定,原生殖细胞可能是一种很好的研究对象。B.C.Wentworth等(1989)提出将鸟类原生殖细胞作为一种有用的基因转移工具。他用采用紫外线照射早期鸡胚的生殖新月区域或以二甲磺酸丁酯注入鸡胚杀死原生殖细胞以及用雄鸡和雌鹌鹑交配的方法获得不育的胚胎,然后将所需研究的原生殖细胞注入不育胚胎来研究其发育和子代的情况。他们将显性的野生鹌鹑的原生殖细胞作为施主注入不育的隐性白羽鹌鹑胚胎受体中,得到两个白色雌性的和一个白色雄性的子代,然后分别再与隐性白羽鹌鹑交配,结果在其子代中获得白羽的和体披野生型羽毛的杂合性子代。J.N.Petitte等(1990)将芦花洛克鸡胚Ⅹ期胚盘注入白来抗鸡胚胎的胚下腔,53个实验中获得6个羽毛表现型为嵌合体胚胎,其中一个孵化为雄性,他们把它与芦花洛克母鸡交配,其目的为测定施主细胞并入宿主睾丸中达到何种程度,在获得719个后代中有两个表现型为芦花洛克,表明施主细胞中的原生殖细胞已并入宿主胚胎的睾丸中,从其血液和精子DNA指纹图像来看不同于来抗鸡。从嵌合体血液DNA指纹图像来看仅为芦花洛克所有。因此,实验结果表明分离芦花洛克X期胚盘细胞提供了来自神经嵴的色素细胞、红血球和生殖细胞。所以,他们认为这种移植胚盘细胞技术可能有助于禽类转基因以及通过体外培养建立多潜能干细胞的研究。【参考文献】:1 Eyal-Giladi H,et al.J Embryo Exp Morph.1981,65:1392 张天荫.细胞生物学杂志,1987,4:1453 张天荫.原生殖细胞(丁汉波等主编·发育生物学).北京:高等教育出版社,19874 Urven E,et al.Anat Rec.,1989,224:245 Wentworth B C,et al.Poultry Sci.1989,68:7996 Loveless W,et al.Development,1990,108:977 刘荣秀.动物学报,1990,36:1148 Petitle J N,et al.Development,1990,108:1859 Godin I,et al.Developnent,1990,108:35710 Nakamura m,et al.Anat Rec,1991,229:109(山东大学张天荫教授撰)