【细胞分类学】
拼译:cytotaxonomy
又称核型系统学。细胞水平分类性状识别、描述分类阶元,并探讨其亲缘关系的学科。主要研究对象是细胞核染色体,即有丝分裂和减数分裂染色体的数目、形态、结构及行为等,研究的是核型,它可分为常规核型和分化核型两个类型,后者又称带型。细胞分类学的基础是在特定的阶元中的整个生命周期、各个组织器官显示出相对稳定的核型,和染色体上基因的位置效应。细胞分类学是细胞遗传学与分类学相互渗透形成的交叉学科,为分类学走向自然分类增加一个重要层次。50年代后,由于低渗、秋水仙碱(素)(纺缍体抑制剂)、PHA(植物血球凝集素)、空气干燥和细胞培养技术的应用,特别是70年代后,带型技术的发展和原位分子杂交技术的应用,使得染色体研究有了很大的发展,各类生物的核带型报道大量涌现,还有专门期刊出版。
体细胞二倍体(2n)染色体是由2个亲本的同源染色体组成的,生殖细胞为单倍体(n),比正常二倍体染色体数目多余染色体,称为超数染色体,或称B染色体。核型中与性别分化相关的称为性染色体,其余的通称常染色体。染色体是由染色质组成的,间期呈现密状者叫异染色质,其余的为常染色质。根据染色体大小和有无明显的着丝点,可以分为大染色体和微小染色体2种类型。大染色体根据着丝点的位(弥漫性着丝点无固定位置)分为(1)中部着丝点(M)、(2)亚中部着丝点(SM)、(3)端部着丝点(T)、(4)亚端部着丝点(ST)等4种类型,在哺乳动物中还用近端着丝点(A)染色体的名称。其类型通常用臂比值(长臂/短臂)来划分(Levan等,1964):臂比值1.0~1.69为M,1.70~2.99为SM,3.0~6.99为ST≥7.0为T,A介于ST和T之间。着丝点把染色体分为2个臂,通常有长臂(q)和短臂(p)之分。一般M和SM计2个臂,T计1个臂,ST和A计臂尚不统一,或1个或2个臂,臂数(NF或AN)是重要的核型参数。另外,次缢痕(h)的数目和位置、随体(Sat.)、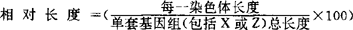 亦是核型参数。如猕猴2n=42(16+24SM)+X(M)+Y(ST),NF=84,20Ph.,峨眉髭蟾2n=26(24m+2sm),NF=52,6qh;昆明龙蜥2n=34(12V+22m),NF=46(爬行动物有鳞目有微小染色体,以m表示,双臂染色体Vv表示,单臂染色体以I表示)。对于减数分裂染色体,通常以其二价体的数目、形状、交叉数目和位置,是否出现多价体等为研究内容。近来,不少学者把前期偶线期——粗线期存在的非永久性核内细胞器——联合复合体(简称SC)作为重要的研究内容,即SC核型分析。由于染色体较长,便于测量,可得到比有系分裂更精确的核型数据。重要的是它可用于观察和研究同源染色体的联合、交换、分离、倒位和易位等结构变化和性染色体的来源等。受某些昆虫的多线染色体带纹的启示,以染色体DNA碱基对非随机排列为假设,1969年Caspersson,首先用合成荧光染料成功地显示了染色体的Q-带后,发展了一系列显示染色体结构的显带技术。常用的有以下几种:G-带是用碱,酶、尿素等处理染色体,使其呈现深浅相间的带纹;以80℃左右的磷酸缓冲液处理可得与G-带相反的带纹,称R-带;通过同步处理获得较长的前中期或后前期染色体,可显示比G-带更清晰更多的带纹,即高分辨G-带。C-带可显示着丝点和其他异染色质区域;银染技术与NOR(核仁组织者区域)、具有活性的28S+18S rRNA基因、次缢痕相关。BrdU渗入使染色体出现DNA复制类型的分化染色,形成复制带。还有碱基特异性荧光带,Hoechst 33258,阿的平、吖啶黄、Distamycin A等为AT-特异性染料;色霉素、光神霉素、放线霉素D等为GC-特异性染料。近年来,有了可对C-带及异染色质区域异质性作差异研究的内切酶带。一般认为带型的产生是染色体经过预处理后,由于核蛋白的选择性提取的产物,是染色体的DNA、蛋白质和染料三者相互作用的结果。对于各类生物大量的核带型研究,表明细胞分类有以下几个意义。核带型的趋异以识别类群;研究阶元分化和物种形成。相同等级内分类阶元间,例如门内纲间、纲内目间、目内科间……以至种内亚种间,亚种内居群间表现出不同的核带型,借此,以检索和鉴别各别分类阶元。以脊椎动物为例,哺乳纲除原兽亚纲有少量微小染色体外,均为大染色体,而鸟纲及爬行纲的核型则有大量的微小染色体;七鳃鳗目有较多(2n>100)染色体,且多为T染色体,而盲鳗目则有数目较少的双臂染色体。在有尾目中,小鲵科含有2n>46,且有许多微小染色体,而蝾螈科和无肺螈科则为20和26~28;人科与猩猩科之间Ag-NORs的分布和数目也有很大的差异。眼镜鳄属与鳄鱼属相比,前者2n=42,后者2n=30-34。棘蛙属内不同种间呈现差异,双团棘胸蛙,棘肛蛙和棘胸蛙的2n分别为64、40和26;麂属内赤麂2n=6(♀),7(♂),而小麂2n=46。最近,在云南高黎贡山发现一新种——贡山麂,2n=8(♀),9(♀)。而原误定为费氏麂其2n=14(♀),15(♂),二者间的核型差异纠正了原来的误定。腹斑掌突蟾与高山掌突蟾的2n分别是22和24,Ag-NOR的位置也不同。黑鹿不同亚种间核型有差异,印度黑鹿2n=58(12M+44A+XY),而云南黑鹿2n=62(8M+52A+XY)。在一些广布种较多存在着不同居群、生态宗间的核型差异,如鲑鳟鱼类中就有种内核型多态现象。阶元间的核型分化,查明其间的染色演化机制和方向,以建立系统发育图。若阶元间的分异时间是已知的,那么就可以定量研究每一个重组的时间。染色体演化与同域物种形成有密切关系。阶元间的核型相似用以探讨共同起源和祖先型,追溯同源性:爬行纲与鸟纲有数目不等的微小染色体,大染色体的G-带相似,人类与猩猩之间G-带相似,硬骨鱼纲许多目核型都由48个T组成,晰蜴类各科都不2n=36(12V+24m)蛇类都有2n=36(16V+20m)等,表明它们的共同起源和同源性。灵长类中从倭狐猴出发,用G-带可以追溯人类各个染色体的来源。核带型研究表明家鼠属中黑家鼠具有原始核型。核型的分化是通过染色体重组实现的,常见的是:(1)罗伯逊易位,即着丝点分离和融合,
亦是核型参数。如猕猴2n=42(16+24SM)+X(M)+Y(ST),NF=84,20Ph.,峨眉髭蟾2n=26(24m+2sm),NF=52,6qh;昆明龙蜥2n=34(12V+22m),NF=46(爬行动物有鳞目有微小染色体,以m表示,双臂染色体Vv表示,单臂染色体以I表示)。对于减数分裂染色体,通常以其二价体的数目、形状、交叉数目和位置,是否出现多价体等为研究内容。近来,不少学者把前期偶线期——粗线期存在的非永久性核内细胞器——联合复合体(简称SC)作为重要的研究内容,即SC核型分析。由于染色体较长,便于测量,可得到比有系分裂更精确的核型数据。重要的是它可用于观察和研究同源染色体的联合、交换、分离、倒位和易位等结构变化和性染色体的来源等。受某些昆虫的多线染色体带纹的启示,以染色体DNA碱基对非随机排列为假设,1969年Caspersson,首先用合成荧光染料成功地显示了染色体的Q-带后,发展了一系列显示染色体结构的显带技术。常用的有以下几种:G-带是用碱,酶、尿素等处理染色体,使其呈现深浅相间的带纹;以80℃左右的磷酸缓冲液处理可得与G-带相反的带纹,称R-带;通过同步处理获得较长的前中期或后前期染色体,可显示比G-带更清晰更多的带纹,即高分辨G-带。C-带可显示着丝点和其他异染色质区域;银染技术与NOR(核仁组织者区域)、具有活性的28S+18S rRNA基因、次缢痕相关。BrdU渗入使染色体出现DNA复制类型的分化染色,形成复制带。还有碱基特异性荧光带,Hoechst 33258,阿的平、吖啶黄、Distamycin A等为AT-特异性染料;色霉素、光神霉素、放线霉素D等为GC-特异性染料。近年来,有了可对C-带及异染色质区域异质性作差异研究的内切酶带。一般认为带型的产生是染色体经过预处理后,由于核蛋白的选择性提取的产物,是染色体的DNA、蛋白质和染料三者相互作用的结果。对于各类生物大量的核带型研究,表明细胞分类有以下几个意义。核带型的趋异以识别类群;研究阶元分化和物种形成。相同等级内分类阶元间,例如门内纲间、纲内目间、目内科间……以至种内亚种间,亚种内居群间表现出不同的核带型,借此,以检索和鉴别各别分类阶元。以脊椎动物为例,哺乳纲除原兽亚纲有少量微小染色体外,均为大染色体,而鸟纲及爬行纲的核型则有大量的微小染色体;七鳃鳗目有较多(2n>100)染色体,且多为T染色体,而盲鳗目则有数目较少的双臂染色体。在有尾目中,小鲵科含有2n>46,且有许多微小染色体,而蝾螈科和无肺螈科则为20和26~28;人科与猩猩科之间Ag-NORs的分布和数目也有很大的差异。眼镜鳄属与鳄鱼属相比,前者2n=42,后者2n=30-34。棘蛙属内不同种间呈现差异,双团棘胸蛙,棘肛蛙和棘胸蛙的2n分别为64、40和26;麂属内赤麂2n=6(♀),7(♂),而小麂2n=46。最近,在云南高黎贡山发现一新种——贡山麂,2n=8(♀),9(♀)。而原误定为费氏麂其2n=14(♀),15(♂),二者间的核型差异纠正了原来的误定。腹斑掌突蟾与高山掌突蟾的2n分别是22和24,Ag-NOR的位置也不同。黑鹿不同亚种间核型有差异,印度黑鹿2n=58(12M+44A+XY),而云南黑鹿2n=62(8M+52A+XY)。在一些广布种较多存在着不同居群、生态宗间的核型差异,如鲑鳟鱼类中就有种内核型多态现象。阶元间的核型分化,查明其间的染色演化机制和方向,以建立系统发育图。若阶元间的分异时间是已知的,那么就可以定量研究每一个重组的时间。染色体演化与同域物种形成有密切关系。阶元间的核型相似用以探讨共同起源和祖先型,追溯同源性:爬行纲与鸟纲有数目不等的微小染色体,大染色体的G-带相似,人类与猩猩之间G-带相似,硬骨鱼纲许多目核型都由48个T组成,晰蜴类各科都不2n=36(12V+24m)蛇类都有2n=36(16V+20m)等,表明它们的共同起源和同源性。灵长类中从倭狐猴出发,用G-带可以追溯人类各个染色体的来源。核带型研究表明家鼠属中黑家鼠具有原始核型。核型的分化是通过染色体重组实现的,常见的是:(1)罗伯逊易位,即着丝点分离和融合, ,引起染色体数目的改变;(2)易位和倒位,导致数目、臂数和内部结构排列变化;(3)常染色质转变为异染色质和异染色质的生长;(4)多倍体化,即基因组重复,等等。已有资料表明类群间的染色体演化速度是很不相同的,如哺乳动物较快,无尾两栖类较慢。核型的保守和分化,取决于诸多因素,特别与物种形成的方式和隔离机制相关。核型演化理论已知的有:(1)循环理论(
,引起染色体数目的改变;(2)易位和倒位,导致数目、臂数和内部结构排列变化;(3)常染色质转变为异染色质和异染色质的生长;(4)多倍体化,即基因组重复,等等。已有资料表明类群间的染色体演化速度是很不相同的,如哺乳动物较快,无尾两栖类较慢。核型的保守和分化,取决于诸多因素,特别与物种形成的方式和隔离机制相关。核型演化理论已知的有:(1)循环理论( )iMAI,1978);(2)隧道模式理论(BicKham & Baker,1979)。核型作为一类性状,也有演化方向和极性(Polarity)问题,在多数情况下,微小染色体逐渐消失,单臂染色体融合为双臂染色体,染色体数目由多变少。但是也有不少的类群中有相反的过程。因此,核型极性的确定要综合形态学,地理分布,古生物学资料及其他进行分析。同样,核型也有平行演化和趋同演化现象。核型作为一种分类性状,同样应遵循后验加权的原则。细胞分类学将随着细胞遗传学和生物分类学的发展和进步,为提高生物分类的预见性作出更大的贡献。但它不可能也不应该回答生物演化和物种形成中的全部问题,但却是一个十分重要的组成成分。【参考文献】:1 Imai H T.J Theor Biol,1978,71∶619~6372 Atchley W R,Woodruff D.Evolution and Speciation,Cambridge Univ Press London,19813 施立明.动物学研究,1984,5(1)(增刊):1~124 King M.Syst Zool,1985,34(1)∶69~755 李树深.生物学通报,1986,8∶1~46 Schmid M,et al.Chromosoma(Berl.)1988,96(4)∶283~290
)iMAI,1978);(2)隧道模式理论(BicKham & Baker,1979)。核型作为一类性状,也有演化方向和极性(Polarity)问题,在多数情况下,微小染色体逐渐消失,单臂染色体融合为双臂染色体,染色体数目由多变少。但是也有不少的类群中有相反的过程。因此,核型极性的确定要综合形态学,地理分布,古生物学资料及其他进行分析。同样,核型也有平行演化和趋同演化现象。核型作为一种分类性状,同样应遵循后验加权的原则。细胞分类学将随着细胞遗传学和生物分类学的发展和进步,为提高生物分类的预见性作出更大的贡献。但它不可能也不应该回答生物演化和物种形成中的全部问题,但却是一个十分重要的组成成分。【参考文献】:1 Imai H T.J Theor Biol,1978,71∶619~6372 Atchley W R,Woodruff D.Evolution and Speciation,Cambridge Univ Press London,19813 施立明.动物学研究,1984,5(1)(增刊):1~124 King M.Syst Zool,1985,34(1)∶69~755 李树深.生物学通报,1986,8∶1~46 Schmid M,et al.Chromosoma(Berl.)1988,96(4)∶283~290(云南大学李树深教授撰)