【喜马拉雅旱獭研究】
拼译:study on marmota hirnalayana
喜马拉雅旱獭又名哈拉、雪猪、曲娃(藏名译音),体形肥硕粗大,体长超过400mm,有的可达650mm;后足长为80mm左右,尾长超过头躯长的1/3,耳高20~25mm;四肢短而粗,前足4指,后足5趾、指端具爪,爪发达适应于掘土,雌兽乳头5对或6对;自鼻端经两眼眉间到两耳前方之间有似三角形的黑色毛区,即“黑三角”,此“黑三角”愈近鼻端愈窄,色调愈黑;嘴四周为黄白色、淡棕黄色或桔黄色;眼眶黑色;面部两颊到耳外侧基部呈淡黄褐色或浅棕黄色,明显有别于“黑三角”;耳壳呈深棕黄色或深黄色;颈背和体背部同色,呈沙黄色、棕黄色或草黄色;毛基为石板色,中段草黄色或淡黄色,毛尖黑色;背部至臀部黑色毛尖多而显著,常形成不规则的黑色细斑纹;体侧黑色毛尖显然较少;体腹面毛稀,毛色呈灰黄色、淡棕色或草黄色;肛门四周和外阴周围呈深棕色或深棕黄色毛区;四肢或足上面呈淡棕黄色或沙黄色,下面与体腹面同色;足掌和爪黑色;尾巴背面毛色与体背相同,毛端约1/4为黑色或黑褐色;尾腹面近基部1/2为棕黄色或褐黄色,端部1/2为黑褐色;毛色随年龄、地区不同而变异。幼体毛色多较成体为暗。有少数白化个体。
头骨粗壮结实,略似三角形。鼻骨较宽而长,前端略超过门齿,后端超过前颌骨后缘,入额骨前端。眶上突发达,向下外方微弯。眶间区凹陷较浅而平坦。颧弓后部明显扩涨,鳞骨前下缘的眶后突起甚小。矢状脊明显,在顶骨长度1/3处的前方分叉,而与眶后突接联呈丫型,幼体头骨无矢状骨脊,脑颅平滑。枕骨大孔前缘呈半椭圆形。腭弓狭长,其后缘超过前颌骨的后缘。腭孔较中国其它诸种旱獭的腭孔长,长8.1~9.1mm。下颌骨的喙状突后缘近乎垂直,不显著向后弯曲,喙状突与关节突之间的切迹深而较窄。牙齿的齿式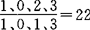 ,上门齿大,前面无沟。上颌第3前臼齿显著,呈圆柱形。上颌前第4前臼齿较大,并臼齿化。上颌臼齿前两枚几乎等大,第3臼齿较大,均具发育较好的原尖、前尖和后尖,且沟深脊显,下颌第4前臼齿之原小尖及下原尖发达。喜马拉雅旱獭的寿命在8a以上,其中1龄组为当年幼体,2龄组为第2年的亚成体,3~6龄为第3~6a的成体,第7~8龄组为第7~8a的老体。生态 喜马拉雅旱獭是青藏高原广泛栖息的动物,它的数量不因草甸草原上植被群落的不同而发生显著变化,主要受地形的影响,山麓平原和山地阳坡下缘是喜马拉雅旱獭集聚的高密度区,阶地山坡和河谷、沟壁为中等密度集聚区,其他地区均为少数或没有,这是最一般的规律。喜马拉雅旱獭营家族生活,通常数个家族形成一个群聚,群聚中个体活动及取食范围互有通达,个体接触密切。群聚中常有个体迁出迁入现象。旱獭的洞穴由主洞(越冬洞)、副洞(夏洞)及临时洞(避敌洞)组成。主洞构造复杂,洞深,洞口较多,内有窝巢,巢内垫有干草。副洞构造较浅而简单,洞口1~2个。临时洞为逃避敌害之用,其构造简单,多在巢域的外围。喜马拉雅旱獭是冬眠动物,整个冬眠期约140d,每年3月底陆续出蛰,10月中旬陆续入蛰。地面活动时间约7个多月,各地出入蛰的时间随各地物候期而有所不同。在室内饲养条件下,旱獭仍有冬眠期,但时间要相对短得多,约110d左右。地面活动范围以6月份最大,9月最小,雄体大于雌体,成体大于幼体,最远可距居住洞口130m左右。旱獭具有一定强度的迁移性,37%属于短距离的或远距离的迁移,其中短距离迁移占20%;远距离迁移占17%,最远在1~2a内可迁移至3000m。旱獭广泛栖息于青藏高原的嵩草草甸草原及针茅草草原,主要分布在东经75°~105°、北纬27°~39°。国外见于喜马拉雅山及喀喇昆化山南坡的克什米尔、尼泊尔、不丹和印度北部等地。国内广泛分布于青藏高原、甘肃祁连山地、阿尔金山及川西北、滇西北等地。垂直分布范围一般多在海拔2800~4000m之间,最低2500m,最高5200m(唐古拉山)。从嵩草草甸草原到针茅草草原带均有分布,分布形式随地形不同而异,在山地多呈带状分布,在平坦地区多呈岛状栖息或弥漫式的栖息,存在个别小片孤立的栖息洞群。食性和食量 野栖喜马拉雅旱獭以沙草科的、禾本科的植物叶、茎及豆科植物的花为食,且有季节变化。春季以青干草为主,日采食量平均350g,夏季以沙草的与禾本科嫩叶为食,日采食量平均500g左右,秋季食沙草科的与禾本科茎叶外和种子,大量采食豆科植物的花(如兰花剌豆),日采食量500g左右。此时是喜马拉雅旱獭增肥最旺时期(解剖成体腹腔内白色脂肪约500g左右,背部与会阴部的白色脂肪约750g左右)。喜马拉雅旱獭在室内饲养条件下表现为杂食性。繁殖特征 旱獭为1年繁殖1次的啮齿动物,3岁始达性成熟,雌性成獭每年仅有一半参加繁殖,妊娠期32~34d。喜马拉雅旱獭种群雌雄性比径卡方检验基本符合1∶1的关系。动物种群繁殖力的又一重要指标取决于性成熟的成年个体占整个种群的比重。据1982~1984年对青海湖东北岸对的调查,达性成熟的旱獭占整个种群的平均值为50.67%。喜马拉雅旱獭的种群繁殖力与种群密度有关,当种群密度大幅度下降时其繁殖力即相应提高,参加繁殖的雌性成獭占具有繁殖力的雌性成熟群体的百分数可高达82.5%,产仔数也可以由年平均数4.71只增至年平均6.93只,说明旱獭密度下降时产仔数明显增多,说明喜马拉雅旱獭动物种群本身存在一种自动调节能力,当种群密度离开平均水平上升或下降时,就有一种使种群重返原来水平的倾向,并且种群数量上升或下降越剧烈,这种相反的倾向也就越明显;换言之,它是随着密度而变化的,是受密度制约的,可以把这种作用称之为反馈。喜马拉雅旱獭繁殖力较弱,每100只雌性成獭繁殖量为255.67,即繁殖季节过后,旱獭种群数量只能增加雌性成獭的2.55倍;但数量大幅度下降时,在其种群内因作用下将明显增加,最高可增加雌性成獭的5.65倍,与小型啮齿动物相比,旱獭种群数量增加的幅度是有限的。喜马拉雅旱獭不管数量变动如何,种群年龄组成不变,性别关系不变,成獭占种群的1/2左右。旱獭9、10月份进入冬眠期,3、4月出蛰,在营地上活动时期的生态习性是变化的,这些变化反应其生理特点,生态期大致可归纳为:(1)出蛰期。旱獭出蛰时间各地不一,随物候期而异,一般为3月下旬至4月中旬,出蛰期约为半个月。刚出蛰活动范围小,离洞较近,仅在中午出洞晒太阳,很少觅食。(2)繁殖期。随着旱獭活动增强即进入繁殖期,平均每天出入洞达10多次,串洞、追逐,吃食时间少。(3)哺育期。仔獭于6月上旬始出生,活动少;为保护幼兽成兽守望增多;仔獭出生有很大一部分不能成活。(4)育肥期。自7月至9月,早獭活动增多,觅食时间增长,幼兽活跃,取食频繁。为觅食方便,成獭可离开主洞,自挖住洞,但距家族主洞均不太远。此期旱獭渐肥,脂肪积累越来越多。(5)入蛰前期。9月以后气候渐冷,植物枯萎,旱獭家族集中于衔草、清理冬眠洞,出入洞频繁。随着气温持续下降,9月底10月初即堵洞,进入蛰眠期。遗传及生理变化 旱獭这一野生哺乳动物细胞遗传特征,用雌雄各6只样本外周血淋巴细胞培养方法,对染色体核型进行研究,结果染色体数目为2n=38,常染色体中有24个中着丝粒染色体和亚中着丝粒染色体,12个端着丝粒染色体。X为中着丝粒染色体,Y为端着丝粒染色体。另一个涉及到遗传背景的问题就是生理功能的遗传基础以及种群遗传。其同功酶和乳酸脱氢酶的应答,用聚丙烯酰胺凝胶盘电泳,对冬眠期的和非冬眠期的喜马拉雅旱獭心、肾、肝和骨骼肌4种组织的乳酸脱氢酶、同功酶带谱分布及酶活力进行研究,结果不同组织的酶谱明显不同,且有多亚带和多态现象,冬眠期各组织的总酶活力高于非冬眠期。喜马拉雅旱獭是耐低氧动物,采用封闭式流体压力呼吸器(Coreeki,1975)测试,当环境温度在15C时,旱獭的耗氧量是2.24ml。喜马拉雅旱獭的一些生理指标测定,用3雌2雄5匹成獭测得均值如下:体重3.35kg,代谢体重2.47kg,体温26.1℃,心率每分钟160次,呼吸频率每分钟21次,每分钟耗氧量0.027L,每分钟产生CO20.0212L,呼吸商0.75,每小时产热量是33J,其基础代谢量每小时千克体重是13.7J。喜马拉雅旱獭血相正常值,血红蛋白103~207g/L,RBC(2.78~5.67)×1012/L,WBC(2.6~12.718)×109/L,中性WBC13.6%~79.2%,淋巴细胞18.0%~86.6%,单核细胞0%~2.7%,嗜酸性白细胞0%~2.3%,嗜碱性WBC0%~0%,以上数值是在玉树地区从100匹成年旱獭现场测得的。经济意义 旱獭肉、骨和油脂均可入药,在藏医中谓其头盖骨(烧成灰,研细)能治水肿;鲜肉和肉干能去寒气,治妇科病;胆汁(胆囊凉干去膜,研细)可治食物中毒、药物中毒及外伤生肌;油脂煎制能去寒气。旱獭皮经加工染色后适制皮领、皮帽和裘衣。1989年,喜马拉雅旱獭在青海省实验动物中心人工驯化繁育成功,初步建立起人工种群,向实验动物种群进化。【参考文献】:1 黄孝龙.兽类学报,1984,5(4):10~162 黄孝龙.兽类学报,1986,6(4):307~3113 梁杰荣.高原生物学集刊,1986,5:91~974 黄孝龙.动物学杂志,1986,21(4):29~315 王懋钦.兽类学报,1989,9(3):173~1756 刘国富.遗传学报,1990,17(2):130~135
,上门齿大,前面无沟。上颌第3前臼齿显著,呈圆柱形。上颌前第4前臼齿较大,并臼齿化。上颌臼齿前两枚几乎等大,第3臼齿较大,均具发育较好的原尖、前尖和后尖,且沟深脊显,下颌第4前臼齿之原小尖及下原尖发达。喜马拉雅旱獭的寿命在8a以上,其中1龄组为当年幼体,2龄组为第2年的亚成体,3~6龄为第3~6a的成体,第7~8龄组为第7~8a的老体。生态 喜马拉雅旱獭是青藏高原广泛栖息的动物,它的数量不因草甸草原上植被群落的不同而发生显著变化,主要受地形的影响,山麓平原和山地阳坡下缘是喜马拉雅旱獭集聚的高密度区,阶地山坡和河谷、沟壁为中等密度集聚区,其他地区均为少数或没有,这是最一般的规律。喜马拉雅旱獭营家族生活,通常数个家族形成一个群聚,群聚中个体活动及取食范围互有通达,个体接触密切。群聚中常有个体迁出迁入现象。旱獭的洞穴由主洞(越冬洞)、副洞(夏洞)及临时洞(避敌洞)组成。主洞构造复杂,洞深,洞口较多,内有窝巢,巢内垫有干草。副洞构造较浅而简单,洞口1~2个。临时洞为逃避敌害之用,其构造简单,多在巢域的外围。喜马拉雅旱獭是冬眠动物,整个冬眠期约140d,每年3月底陆续出蛰,10月中旬陆续入蛰。地面活动时间约7个多月,各地出入蛰的时间随各地物候期而有所不同。在室内饲养条件下,旱獭仍有冬眠期,但时间要相对短得多,约110d左右。地面活动范围以6月份最大,9月最小,雄体大于雌体,成体大于幼体,最远可距居住洞口130m左右。旱獭具有一定强度的迁移性,37%属于短距离的或远距离的迁移,其中短距离迁移占20%;远距离迁移占17%,最远在1~2a内可迁移至3000m。旱獭广泛栖息于青藏高原的嵩草草甸草原及针茅草草原,主要分布在东经75°~105°、北纬27°~39°。国外见于喜马拉雅山及喀喇昆化山南坡的克什米尔、尼泊尔、不丹和印度北部等地。国内广泛分布于青藏高原、甘肃祁连山地、阿尔金山及川西北、滇西北等地。垂直分布范围一般多在海拔2800~4000m之间,最低2500m,最高5200m(唐古拉山)。从嵩草草甸草原到针茅草草原带均有分布,分布形式随地形不同而异,在山地多呈带状分布,在平坦地区多呈岛状栖息或弥漫式的栖息,存在个别小片孤立的栖息洞群。食性和食量 野栖喜马拉雅旱獭以沙草科的、禾本科的植物叶、茎及豆科植物的花为食,且有季节变化。春季以青干草为主,日采食量平均350g,夏季以沙草的与禾本科嫩叶为食,日采食量平均500g左右,秋季食沙草科的与禾本科茎叶外和种子,大量采食豆科植物的花(如兰花剌豆),日采食量500g左右。此时是喜马拉雅旱獭增肥最旺时期(解剖成体腹腔内白色脂肪约500g左右,背部与会阴部的白色脂肪约750g左右)。喜马拉雅旱獭在室内饲养条件下表现为杂食性。繁殖特征 旱獭为1年繁殖1次的啮齿动物,3岁始达性成熟,雌性成獭每年仅有一半参加繁殖,妊娠期32~34d。喜马拉雅旱獭种群雌雄性比径卡方检验基本符合1∶1的关系。动物种群繁殖力的又一重要指标取决于性成熟的成年个体占整个种群的比重。据1982~1984年对青海湖东北岸对的调查,达性成熟的旱獭占整个种群的平均值为50.67%。喜马拉雅旱獭的种群繁殖力与种群密度有关,当种群密度大幅度下降时其繁殖力即相应提高,参加繁殖的雌性成獭占具有繁殖力的雌性成熟群体的百分数可高达82.5%,产仔数也可以由年平均数4.71只增至年平均6.93只,说明旱獭密度下降时产仔数明显增多,说明喜马拉雅旱獭动物种群本身存在一种自动调节能力,当种群密度离开平均水平上升或下降时,就有一种使种群重返原来水平的倾向,并且种群数量上升或下降越剧烈,这种相反的倾向也就越明显;换言之,它是随着密度而变化的,是受密度制约的,可以把这种作用称之为反馈。喜马拉雅旱獭繁殖力较弱,每100只雌性成獭繁殖量为255.67,即繁殖季节过后,旱獭种群数量只能增加雌性成獭的2.55倍;但数量大幅度下降时,在其种群内因作用下将明显增加,最高可增加雌性成獭的5.65倍,与小型啮齿动物相比,旱獭种群数量增加的幅度是有限的。喜马拉雅旱獭不管数量变动如何,种群年龄组成不变,性别关系不变,成獭占种群的1/2左右。旱獭9、10月份进入冬眠期,3、4月出蛰,在营地上活动时期的生态习性是变化的,这些变化反应其生理特点,生态期大致可归纳为:(1)出蛰期。旱獭出蛰时间各地不一,随物候期而异,一般为3月下旬至4月中旬,出蛰期约为半个月。刚出蛰活动范围小,离洞较近,仅在中午出洞晒太阳,很少觅食。(2)繁殖期。随着旱獭活动增强即进入繁殖期,平均每天出入洞达10多次,串洞、追逐,吃食时间少。(3)哺育期。仔獭于6月上旬始出生,活动少;为保护幼兽成兽守望增多;仔獭出生有很大一部分不能成活。(4)育肥期。自7月至9月,早獭活动增多,觅食时间增长,幼兽活跃,取食频繁。为觅食方便,成獭可离开主洞,自挖住洞,但距家族主洞均不太远。此期旱獭渐肥,脂肪积累越来越多。(5)入蛰前期。9月以后气候渐冷,植物枯萎,旱獭家族集中于衔草、清理冬眠洞,出入洞频繁。随着气温持续下降,9月底10月初即堵洞,进入蛰眠期。遗传及生理变化 旱獭这一野生哺乳动物细胞遗传特征,用雌雄各6只样本外周血淋巴细胞培养方法,对染色体核型进行研究,结果染色体数目为2n=38,常染色体中有24个中着丝粒染色体和亚中着丝粒染色体,12个端着丝粒染色体。X为中着丝粒染色体,Y为端着丝粒染色体。另一个涉及到遗传背景的问题就是生理功能的遗传基础以及种群遗传。其同功酶和乳酸脱氢酶的应答,用聚丙烯酰胺凝胶盘电泳,对冬眠期的和非冬眠期的喜马拉雅旱獭心、肾、肝和骨骼肌4种组织的乳酸脱氢酶、同功酶带谱分布及酶活力进行研究,结果不同组织的酶谱明显不同,且有多亚带和多态现象,冬眠期各组织的总酶活力高于非冬眠期。喜马拉雅旱獭是耐低氧动物,采用封闭式流体压力呼吸器(Coreeki,1975)测试,当环境温度在15C时,旱獭的耗氧量是2.24ml。喜马拉雅旱獭的一些生理指标测定,用3雌2雄5匹成獭测得均值如下:体重3.35kg,代谢体重2.47kg,体温26.1℃,心率每分钟160次,呼吸频率每分钟21次,每分钟耗氧量0.027L,每分钟产生CO20.0212L,呼吸商0.75,每小时产热量是33J,其基础代谢量每小时千克体重是13.7J。喜马拉雅旱獭血相正常值,血红蛋白103~207g/L,RBC(2.78~5.67)×1012/L,WBC(2.6~12.718)×109/L,中性WBC13.6%~79.2%,淋巴细胞18.0%~86.6%,单核细胞0%~2.7%,嗜酸性白细胞0%~2.3%,嗜碱性WBC0%~0%,以上数值是在玉树地区从100匹成年旱獭现场测得的。经济意义 旱獭肉、骨和油脂均可入药,在藏医中谓其头盖骨(烧成灰,研细)能治水肿;鲜肉和肉干能去寒气,治妇科病;胆汁(胆囊凉干去膜,研细)可治食物中毒、药物中毒及外伤生肌;油脂煎制能去寒气。旱獭皮经加工染色后适制皮领、皮帽和裘衣。1989年,喜马拉雅旱獭在青海省实验动物中心人工驯化繁育成功,初步建立起人工种群,向实验动物种群进化。【参考文献】:1 黄孝龙.兽类学报,1984,5(4):10~162 黄孝龙.兽类学报,1986,6(4):307~3113 梁杰荣.高原生物学集刊,1986,5:91~974 黄孝龙.动物学杂志,1986,21(4):29~315 王懋钦.兽类学报,1989,9(3):173~1756 刘国富.遗传学报,1990,17(2):130~135(青海省地方病防治研究所黄孝龙副主任医师撰;吴祥林审)