【柑桔类病毒及类病毒病】
拼译:citrus viroid and viroid disease
类病毒是目前已知的最小的致病因子。它是一种无蛋白质外壳的环状单链RNA分子,只含有246~375个核苷酸;由于分子内存在着广泛的碱基对,而形成类似于棒状的二级结构。至今仅在植物中发现它们,它能对许多栽培作物的生长产生显著影响。类病毒携带的遗传信息量极小,其RNA的长度不足以编码一个蛋白质,它的增殖可能完全依赖于寄主中尚不清楚的复制机构。在柑桔上,柑桔裂皮类病毒(CEVd)是继马铃薯纺锤型块茎类病毒(PSTVd)之后发现的第2种类病毒,它们的发现开辟了病毒学的一个新领域——亚病毒。除CEVd外还存在其他几种柑桔类病毒(CVd),它们组成单一,结构简单,寄主范围相近,引起症状类型相似,是研究致病机理、病害防治及生命起源和演化的有利材料,亦是研究分子结构和生物功能的良好体系。 1950年,本顿(R.J.Benton)等报道了以枳、枳橙和 檬作砧木的柑桔树砧木部树皮纵向开裂或翘裂、植株矮化等柑桔裂皮病的田间症状及传染性。同年,蔡尔兹(J.F.L.Childs)等又报道了柑桔木质陷孔病的传染性和在桔柚上引起的流胶、陷点和木刺等症状。由于当时对植物病原物认识的局限,认为它们可能是由病毒引起的病害,但随后即发现其病原与病毒有很大的差异。1967年,韦瑟斯(L.G.Weathers)等发现了一种CEVd的专一性增殖、鉴别寄主爪哇三七;1971年迪内(T.O.Diener)在马铃薯上发现一种新的小分子RNA植物病原PSTVd后,1972年塞曼斯克(J.S.Semancik)等对从爪畦三七中分离到的柑桔裂皮病病原的热稳定性、苯酚处理的耐性、对核酸酶的敏感性以及电泳迁移率等理化性质的研究证明柑桔裂皮病亦是由一种类病毒引起的病害。长期以来对柑桔木质陷孔病是类病毒病害的怀疑,直到1986年发现了黄瓜和Etrog香橼是木质陷孔病病原的良好增殖寄主,和应用更加灵敏的类病毒分离及诊断技术后才得到塞曼斯克等人的证实。在类病毒的研究中,一些分离和诊断技术的改进和发展对柑桔类病毒病原的认识起到了关键的作用。这些技术包括:(1)双向聚丙烯酰胺凝胶电泳技术能有效地分离寄主及类病毒的线状的和环状的核酸;(2)溴乙淀(EB)或银染色能提高寄主中低浓度类病毒的检测;(3)互补DNA(cDNA)探针技术能区分类病毒之间的同源性;(4)选择性的寄主系统能鉴别不同类病毒的生物学反应和有利于类病毒的增殖、纯化。应用这些技术已发现的CVd共有12种,为了便于研究和比较,杜兰维拉(N.Duran-Vila)等根据它们不同的物理学的和生物学的性质将其划分为5个组(表1)。 檬作砧木的柑桔树砧木部树皮纵向开裂或翘裂、植株矮化等柑桔裂皮病的田间症状及传染性。同年,蔡尔兹(J.F.L.Childs)等又报道了柑桔木质陷孔病的传染性和在桔柚上引起的流胶、陷点和木刺等症状。由于当时对植物病原物认识的局限,认为它们可能是由病毒引起的病害,但随后即发现其病原与病毒有很大的差异。1967年,韦瑟斯(L.G.Weathers)等发现了一种CEVd的专一性增殖、鉴别寄主爪哇三七;1971年迪内(T.O.Diener)在马铃薯上发现一种新的小分子RNA植物病原PSTVd后,1972年塞曼斯克(J.S.Semancik)等对从爪畦三七中分离到的柑桔裂皮病病原的热稳定性、苯酚处理的耐性、对核酸酶的敏感性以及电泳迁移率等理化性质的研究证明柑桔裂皮病亦是由一种类病毒引起的病害。长期以来对柑桔木质陷孔病是类病毒病害的怀疑,直到1986年发现了黄瓜和Etrog香橼是木质陷孔病病原的良好增殖寄主,和应用更加灵敏的类病毒分离及诊断技术后才得到塞曼斯克等人的证实。在类病毒的研究中,一些分离和诊断技术的改进和发展对柑桔类病毒病原的认识起到了关键的作用。这些技术包括:(1)双向聚丙烯酰胺凝胶电泳技术能有效地分离寄主及类病毒的线状的和环状的核酸;(2)溴乙淀(EB)或银染色能提高寄主中低浓度类病毒的检测;(3)互补DNA(cDNA)探针技术能区分类病毒之间的同源性;(4)选择性的寄主系统能鉴别不同类病毒的生物学反应和有利于类病毒的增殖、纯化。应用这些技术已发现的CVd共有12种,为了便于研究和比较,杜兰维拉(N.Duran-Vila)等根据它们不同的物理学的和生物学的性质将其划分为5个组(表1)。表1 柑桔类病毒物理和生物学性质的关系 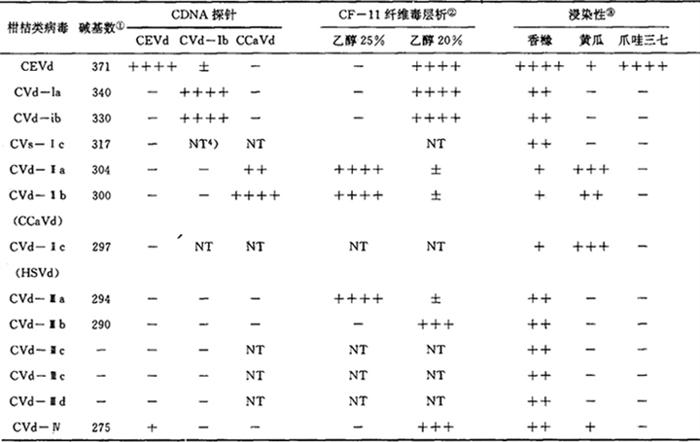
①碱基数是在变性的聚丙烯酰胺凝胶电泳条件下,以CEVd和ASVd作参照测得。②CF-11纤维素洗脱表示类病毒开始洗脱时的乙醇百分含量。③浸染性:++++ =严重症状 + =受感染,无症状。 +++ =中等症状 - =未受感染,无症状。 ++ =轻度症状④NT=未测定 (本表引至杜兰维拉)柑桔中一些新的类病毒的发现,丰富了人们对柑桔裂皮病和木质陷孔病的认识。1964年,卡拉范(E.C.Calavan)等报道了Etrog香橼是鉴定裂皮病的指示植物后,通常认为引起Etrog香橼表现严重、中等和轻度卷叶等症状的分别是CEVd的强、中等和弱株系。然而,这种由于症状类型相以而引起的臆测,没有经过在枳壳等上的进一步确认。1985年施莱默(A.Schlemmer)等发现,似引起Etrog香橼表现中等和轻度卷叶病状的病原不是CEVd,而是一些分子量更小的CVd后,CEVd是柑桔裂皮病的唯一病原的观点受到怀疑。1988年,吉林斯(M.R.Gillings)等对田间枳砧柑桔树感染CVd情况的初步分析表明,田间裂皮病树通常受到几种CVd的复合感染,除CEVd外一些分子量更小的CVd也能引起枳壳发生轻度裂皮和(或)矮化等症状。因此,本顿等人描述的柑桔裂皮病并非是单一病原引起的,实际上它是一些CVd引起的综合症。对木质陷孔病的研究也表明,引起木质陷孔病的病原不只木质陷孔类病毒(CCaVd)一种,CVd-Ⅱc(洒花矮化类病毒HSVd的柑桔株系)亦能引起类似的症状。CVd复合感染,不同CVd引起相似的症状表现以及同一种CVd存在不同株系,是CVd与柑桔裂皮病和木质陷孔病相互关系复杂化的原因。幸运的是,塞曼斯克等人在研究CEVd的理化性质和核酸序列时应用了爪哇三七这种CEVd专一性的寄主,从而避免了其他柑桔类病毒对研究结果的干扰。CEVd引起柑桔树的症状并不都是有害的,CEVd强株系对枳壳砧柑桔树的严重危害只是一个极端的例子。大多数情况下,CVd引起柑桔树的生长变化并不影响到柑桔生产,而有的影响对柑桔生产是有利的。1968年,科恩(M.Cohen)等首先提出了利用CEVd的弱株系进行矮化栽培的实验,为CVd的利用开创了先例。目前,许多国家都在开展这方面的研究工作,但尚未大量应用于田间生产。对CVd病害的防治,主要是应用茎尖培养脱除类病毒,繁育无类病毒苗木的方法。研究一些针对强致病CVd的防治方法,对CVd的利用和严重CVd病害的防治是有利的。1978年中国已有柑桔裂皮病发生的报道,随后赵学源等用Etrog香橼进行了鉴定。1985年,糜克永等从柑桔裂皮病树中分离到CEVd。田间柑桔树中发现其他的柑桔病毒和类似于类病毒RNA已见报道。随着一种新的柑桔小分子RNA病原CEVd的证明,对其结构和核酸序列、复制、致病过程以及与寄主细胞之间的干扰进行了大量研究。CEVd是至今报道的最大的类病毒,它由371个核苷酸组成,具有线状的和环状的两种形式,它们均能引起爪哇三七产生同样的症状。1982年,格罗斯(H.J.Gross)和维斯瓦德(J.E.Visvader)等人对CEVd核酸序列进行了直接RNA或cDNA分析。CEVd的所有株系都包含371个核苷酸,不同株系间有多到27个核苷酸在交换、插入和缺失上的差异。CEVd也存在着大量的田间变异体,变异体的核苷酸数大约在370~375个之间。已报道的田间变异体根据其在番茄上的症状差异被分为CEVd-A和CEVd-B两类,CEVd-A引起番茄表现严重症状,CEVd-B引起轻度症状。1985年,基斯(P.Keese)等在研究CEVd时将类病毒类似于棒状的二级结构划分为5个结构功能区。1986年,维斯瓦德等用CEVd-A和CEVd-B的部分序列构建的嵌合体浸染性cDNA质粒显示了致病区对症状表现程度的控制,和可变区对类病毒浓度的影响。1982年,弗洛里斯(R.Flores)等人在研究CEVd的复制时发现CEVd感染的爪哇三七富含核的制剂中含高浓度的CEVd,而未受CEVd感染的爪哇三七富核制剂能在体外合成CEVd。一种能抑制细胞核中依赖于DNA的PNA聚合酶Ⅱ的抑制剂α-鹅膏亭能抑制CEVd的体外合成,表明CEVd的复制是在核中进行,并需要一种寄主编码的DNA转录酶的催化。CEVd对这种酶的干扰可能是类病毒致病的原因之一。1985年,哈钦斯(C.Hutchins)等对类病毒复制过程的研究提出了类病毒复制的滚环模型,检测到寄主中存在CEVd的互补RNA和多聚体CEVd支持了这一复制假说。类病毒引起寄主的症状反应与寄主细胞的结构、代谢、植物激素平衡的改变有关。塞曼斯克等人对CEVd引起的细胞结构变化的一系列研究发现,CEVd感染细胞后细胞壁中富含羟脯氨酸的糖蛋白和阿拉伯半乳糖残基的增加影响到细胞壁厚度的增加和细胞的伸长,β-1.3葡聚糖相对比例的改变和在细胞壁表面上不均匀的堆积导致细胞壁畸变。CEVd感染的组织中细胞与细胞间连接的改变减小了细胞间的空间。细胞壁的畸变、膜结构的紊乱以及细胞间隙的减小,可能是引起寄主宏观症状的原因。CEVd感染细胞后主要引起细胞中碳水化合物代谢的变化,细胞中核酸的量和质均未有可检测到的任何改变。虽然在CEVd感染的细胞中不存在与CEVd有关的特异性蛋白质产物,但CEVd的感染能增进寄主编码的一些蛋白质的合成。科奈杰罗(V.Conejero)研究组在这方面进行的长期工作证明,其中几种蛋白质的功能,P69具有蛋白酶活性,P32和P34具有几丁质酶的功能。然而CEVd引起的蛋白质量的改变并非是特异的。其它病原物和银离子也能引起类似的改变。CEVd感染寄主后引起植株的矮化显然与改变了寄主中植物激素的平衡有关。弗洛里斯等研究了CEVd感染的爪哇三七中一种生长素类物质的减少影响到根的发生,然而杜兰维拉等发现CEVd感染的番茄组织在各种浓度的IAA下培养,IAA对根的发生并没有促进作用。可能是CEVd感染后引起细胞壁的畸变和组织结构的损害影响到培养组织对生长素的利用。科奈杰罗等研究表明,受CEVd感染的植株中乙烯的增加引起严重矮化、卷叶、退绿、早熟、早衰、少根等症状。向健康的番茄施用一种乙稀的前体(ACC)、乙烯利等能产生和CEVd感染后相似的症状,并且与CEVd一样能引起寄主的某些蛋白质增加。这些实验结果表明,外部因子如类病毒刺激起动了寄主代谢的二级偶联,但目前这些变化的因果关系是不清楚的。愈伤组织和细胞悬浮培养作为一种更加精确、容易控制的研究系统已被广泛用于类病毒的感染、复制和致病机理的研究。林(J.J.Lin)、马顿(L.Marton)和王(M.C.Wang)等人的实验结果表明,类病毒引起的植物组织细胞的变化也能在无组织结构的培养细胞中发现;在一些特定的培养条件下,受CEVd感染的细胞比健康的细胞表现出更强的生长活性和对植物激素、糖的耐性,说明CEVd在一定条件下对细胞的生长是有利的。因此有人认为,类病毒不是致病因子而是调节因子控制细胞生长。已知的CVd在发现它的国家其分布是不同的,其他国家是否存在新的CVd有待调查研究,这些CVd单独感染或复合感染后引起植株的生长变化,是近期研究的一个重要方面。利用构建类病毒的cDNA浸染性质粒和用PCR合成类病毒单体技术研究CEVd和其它的CVd分子结构与功能的关系有着广泛的前景,合成的突变体类病毒,也可能用于控制柑桔有利地生长,对CEVd等复制的研究将在更加精确的细胞核溶解物中进行。在柑桔组织和细胞培养系统中研究类病毒-寄主基因表达-蛋白质(酶)、植物激素-寄主生长变化的相互关系,有利于揭示类病毒致病过程和干扰寄主的具体作用位点,通过该系统也可进行类病毒介入的柑桔组织和细胞抗逆性研究。植物转基因工程为转移特异性切割强致病CVd株系的核酶(Ribozyme)基因于寄主中,防治类病毒病害发生提供了一条新途径,它可能防除强致病类病毒而保留一些对柑桔生长有利的类病毒。利用类病毒进行的柑桔矮化栽培将在统一的操作规程和管理制度下试用于田间生产。【参考文献】:1 Semancik J S,et al.Physical Properties of a minimal infectious RAN(viroid)associated with the exocotis disease.Viro-logy,1975,63,160~1672 Visvader J E,et al.Infectivity and in vitro mutagenesis of monomeric cNA clones of citrus exocortis viroid indcates the site of Procecssing of viroid precursors.Nucleic Acids Res.,1985,13:16,5843~58563 Schlemner A,et al.A unique,infectious RNA associated with citron showing symptoms typical of citrus exocortis viroid.Phytopathology,1985,75:946~9494 Semancik J S.The citrus exocoris disease.Areview 1976~1986.In Proc.10th Conf.IOCV,Riverside,1986.136~1515 Gillings M R,et al.Viroids in Australian citrus.Proceedings of the Sixth International Citrus Congress,1988,3:881~8956 Rivera-Bustamante R F,et al.Properties of a Viroid-rep-licating complex solubilized from nuclei.J.Gen.Virol.,1989,70:10,2707~27167 Garcia F J,et al.C7(P32)and C6(P34)PR proteins induced in tomato leaves by citrus exocortis viroid infection are chtinases.Physiol.Mol.Pathol.,1990,36:3,249~2608 Juarez J,et al.Separation of citars viroids by shoot-tip graftiong in vito.Plant Pathol.,1990,39(3):472~4769 欧阳立,等.中国栽培柑桔中存在类似病毒RNA的证明.植物病理学报,1990,20(2):13410 Rigden J E,et al.In vitro synthesis of an infectious viroid:analysis of the infectivity of monomeric linear CEVd.Virology,1992,186,201~206(中国农业科学院柑桔研究所欧阳立撰) |