【遗传密码】
遗传信息的表达是信息分子核酸指导生成功能分子蛋白质的过程。RNA(DNA)由4种单核苷酸A、C、G、U(A、C、G、T)组成,蛋白质由20种氨基酸组成,信息从前者传递到后者要求核苷酸和氨基酸间存在一定对应关系,由核苷酸编码氨基酸。
1953年沃森(Watson)和克里克(Crick)提出DNA的双螺旋结构模型后,伽莫夫(Gamow)(1954)首先提出由DNA的4种碱基来编码20种氨基酸的遗传密码思想。认为蛋白质在DNA表面上合成,氨基酸直接位于双螺旋的沟中,在4个碱基构成的菱形小孔中置放一个氨基酸。证明44=256种菱形中,在一定约束下正好填充20种氨基酸(金刚石密码)。伽莫夫认为DNA是蛋白质合成模板的观点与实验不符,并且这种密码是交迭4联体,也与后来证明的实际密码是非交迭3联体相矛盾。从基本的信息论考虑便可导出编码一个氨基酸的核苷酸(碱基)数不能小于3。因为一个氨基酸的信息量是4.32比特,而一个核苷酸的信息量只有2比特。碱基点突变的实验证明,一个碱基的变化只会引起一个氨基酸的变化,碱基缺失可为碱基插入的移码突变所补偿。这些实验证明了密码是非交迭的三联体。历史上密码的破译是通过蛋白质的生物合成获得解决的。1961年尼伦伯格(Nirenbberg)和马太埃(Matthaei)及乔乔阿(Ochoa)等两个组开始用人工合成的RNA进行无细胞蛋白质合成。这个无细胞系统的制备通过破坏大肠杆菌细胞得到,它包含核糖体:tRNA,ATP和GTP;以及必需的酶,但不含DNA和mRNA。然后引入人工合成的poly-U,便可观察到苯丙氨酸的合成。如果换成Poly-C,便可观察到脯氨酸。后尼伦伯格将上述实验中的多核苷换成各种已知结构的3-核苷酸(密码子),研究3-核苷-tRNA-氨基酸复合体,检验了约50多种密码子,从而建立了密码子氨基酸的对应关系。1966年柯拉纳(Khorana)进一步将有机化学合成和酶合成方法结合起来,用人工合成约12个单位的寡脱氧核糖核苷酸为模板,在DNA聚合酶作用下,通过重复复制,生成类DNA双螺旋,其两股各包含互补的重复若干次的核苷三联体。再藉助于RNA聚合酶,由类-DNA产生已知顺序的RNA,把它们引入无细胞系统,便得到各种对应的氨基酸。柯拉纳通过人工模拟氨基酸的生物合成,证实了尼伦格伯的实验,从而完全破译了遗传密码。尼伦伯格和柯拉纳的工作获得1968年诺贝尔生理医学奖。遗传密码的重要特点是它的普适性(通用性)。但1979年后发现了一些密码反常,总结如下(wong,1988):
 。这个精确度已大大超过非生物学反应。密码字典的一个重要特点是它的简并性——几个密码子(称为同义密码子)编码同一个氨基酸。普通密码中61个有意义的密码子编码了20个氨基酸。其中AUG除编码甲硫氨酸外,还可与fMet-tRNA结合,作为起始密码。GUG也可作起读之用。UAG、UAA、UGA不能与tRNA结合,它们是无义密码子,作为终止符号。在反常密码中,起始和终止密码常有变化。哺乳类线粒体使用AUU、AUC、AUA为起始密码,终止密码的反常见表。核酸编码氨基酸是通过mRNA上的密码子与tRNA上的反密码子结合而实现的。对应于一种氨基酸,存在一种或几种tRNA,它们一般以第34~36位携带的不同反密码子相区别。理论上tRNA的种类数应在20和61之间。克里克提出摆动说(1966),认为tRNA5′端空间位置不甚固定,能和密码子3′端两种或3种碱基配对。后来证明这种摆动还和反密码子5′端的核苷转变为某些修饰成分(如次黄苷I,I能与C、U、A配对)有关。克里克摆动是密码简并的重要原因。在一个简并多重态中,诸密码子使用频率不同,它受着基因组碱基频率的约束(Bernardi,1985)。人们推测它与基因组中相应tRNA的拷贝数有关(Ikemura,1985)。统计分析也表明它可能与基因表达水平有关,产生高丰度蛋白的高表达基因使用高偏置的密码子(Gouy等,1982;Sharp等,1986)。从已测定一级结构的tRNA的统计分析表明,在其60几个至90几个核苷中,除恒定和半恒定核苷约占
。这个精确度已大大超过非生物学反应。密码字典的一个重要特点是它的简并性——几个密码子(称为同义密码子)编码同一个氨基酸。普通密码中61个有意义的密码子编码了20个氨基酸。其中AUG除编码甲硫氨酸外,还可与fMet-tRNA结合,作为起始密码。GUG也可作起读之用。UAG、UAA、UGA不能与tRNA结合,它们是无义密码子,作为终止符号。在反常密码中,起始和终止密码常有变化。哺乳类线粒体使用AUU、AUC、AUA为起始密码,终止密码的反常见表。核酸编码氨基酸是通过mRNA上的密码子与tRNA上的反密码子结合而实现的。对应于一种氨基酸,存在一种或几种tRNA,它们一般以第34~36位携带的不同反密码子相区别。理论上tRNA的种类数应在20和61之间。克里克提出摆动说(1966),认为tRNA5′端空间位置不甚固定,能和密码子3′端两种或3种碱基配对。后来证明这种摆动还和反密码子5′端的核苷转变为某些修饰成分(如次黄苷I,I能与C、U、A配对)有关。克里克摆动是密码简并的重要原因。在一个简并多重态中,诸密码子使用频率不同,它受着基因组碱基频率的约束(Bernardi,1985)。人们推测它与基因组中相应tRNA的拷贝数有关(Ikemura,1985)。统计分析也表明它可能与基因表达水平有关,产生高丰度蛋白的高表达基因使用高偏置的密码子(Gouy等,1982;Sharp等,1986)。从已测定一级结构的tRNA的统计分析表明,在其60几个至90几个核苷中,除恒定和半恒定核苷约占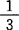 外,某些位点具有特异性,它们有决定该分子选择性地携带某种氨基酸的作用(例如Ala-tRNA的第3位G和第70位U),称为第2遗传密码(de Duve,1988;Hou等,1988)。这方面的工作正在进展。关于遗传密码的物理意义,早期工作已知密码子第2碱基对氨基酸理化性质有决定作用。如把碱基突变分成保守和改变氨基酸疏水性的两类,那么后一类突变是危险的,并已为自然选择所消除。因此密码表的结构有利于前一类突变(Volkenstein,1977)。80年代以后开展的遗传密码逻辑的研究,包括密码的简并规则,简并多重态的分布,密码表的疏水-亲水畴,以及密码的对偶性等。通过引入密码字典突变危险性的概念,从它的极小化可导出普适密码表中各种简并多重态的排列规则及出现频率,并解释亲水和疏水氨基酸在密码表中的畴状结构。从而由稳定性原理导出了普适密码表的各种特征(Luo,1988;1989)。密码的阴阳对偶性的研究揭示了密码逻辑的另一重要方面,由此也可较好解释氨基酸的疏水-亲水分布(Luo,1992)。遗传密码理论研究的另一个焦点是密码的起源和进化,从氨基酸的合成实验和陨石资料都说明Gly,Ala,Ser,Asp,Glu和Val等是最早出现的氨基酸,可能它们首先被编码。有人从线粒体密码推测原始密码,认为原始的tRNA结构简并,没有专一性,密码三联体实际只是“三中读二”,进化朝着减少密码模糊性的方向发展(Jukes,1983)。遗传密码的研究在人类对生命奥秘的探索中占据重要地位,它不仅具有理论意义,而且在蛋白质工程中有重要实践价值。期望通过密码突变来改造氨基酸(包括产生非标准氨基酸)从而改造蛋白质-这可能是蛋白质工程和设计的一个新方向。
外,某些位点具有特异性,它们有决定该分子选择性地携带某种氨基酸的作用(例如Ala-tRNA的第3位G和第70位U),称为第2遗传密码(de Duve,1988;Hou等,1988)。这方面的工作正在进展。关于遗传密码的物理意义,早期工作已知密码子第2碱基对氨基酸理化性质有决定作用。如把碱基突变分成保守和改变氨基酸疏水性的两类,那么后一类突变是危险的,并已为自然选择所消除。因此密码表的结构有利于前一类突变(Volkenstein,1977)。80年代以后开展的遗传密码逻辑的研究,包括密码的简并规则,简并多重态的分布,密码表的疏水-亲水畴,以及密码的对偶性等。通过引入密码字典突变危险性的概念,从它的极小化可导出普适密码表中各种简并多重态的排列规则及出现频率,并解释亲水和疏水氨基酸在密码表中的畴状结构。从而由稳定性原理导出了普适密码表的各种特征(Luo,1988;1989)。密码的阴阳对偶性的研究揭示了密码逻辑的另一重要方面,由此也可较好解释氨基酸的疏水-亲水分布(Luo,1992)。遗传密码理论研究的另一个焦点是密码的起源和进化,从氨基酸的合成实验和陨石资料都说明Gly,Ala,Ser,Asp,Glu和Val等是最早出现的氨基酸,可能它们首先被编码。有人从线粒体密码推测原始密码,认为原始的tRNA结构简并,没有专一性,密码三联体实际只是“三中读二”,进化朝着减少密码模糊性的方向发展(Jukes,1983)。遗传密码的研究在人类对生命奥秘的探索中占据重要地位,它不仅具有理论意义,而且在蛋白质工程中有重要实践价值。期望通过密码突变来改造氨基酸(包括产生非标准氨基酸)从而改造蛋白质-这可能是蛋白质工程和设计的一个新方向。(内蒙古大学博士生导师罗辽复教授撰)