【光敏核不育水稻的育性转换】
光敏核不育水稻是具有雄性育性在一定光温条件下可以在败育与正常可育之间变化的新兴核不育水稻。在杂交水稻研究历史上,野败不育基因的发现使陷入困境的水稻杂种优势利用研究在中国首先突破。但三系法的繁殖制种以及特定的恢保关系给杂交稻的利用带来许多限制。光敏核不育水稻(PGMR)的发现可以简化繁殖过程,克服恢保关系限制,为更加有效地利用水稻杂种优势提供了保贵的遗传工具。 1973年,石明松在湖北沔阳沙湖原种场农垦58水稻田中,发现一株自然不育株。受野败不育材料的利用途径的影响,进行大量测交以寻找保持系,但均未成功。在研究中,发现头季不育的材料割蔸再生可收到些种子,而这些自交种在下年头季种植还可继续保持不育特性。初步认为,诱导其育性转换的主导因子是日照长度,而不育性与当地自然温度的相关性不显著。后来的人工光照试验,证实了光周期在诱导PGMR育性转换中的关键作用。并且得出诱导PGMR光敏农垦58S育性转换的临界光长为13h45min~14h00min;临界光强为50lx;在短日光周期的长暗期中,短时光间断也能起长日诱导不育的作用,诱导PGMR育性转换的关键因素为暗期长度。在PGMR不同的发育时期所进行的光照处理试验中,发现光周期诱导育性转换的敏感时期为幼穗分化的第2次枝梗及小穗原基分化期至花粉母细胞形成期。光周期只有在这一敏感期才具有诱导育性转换的作用。敏感期全为长日条件可诱导全不育;部分处在长日下诱导半不育;全在短日下花粉可接近正常可育。诱导花粉育性转换的作用时期与诱导植株进入生殖生长的敏感期发生在不同时期。从而,提出了PGMR两个光周期反应理论。并对两个光周期反应的诱导条件、生理过程、遗传关系等作了系列探讨,发现两个光周期反应在PGMR的发育过程中属于相对独立的生理遗传过程,可以通过育种手段予以重组。由于在研究育性转换的光周期性过程中,发现相同的处理在不同季节中进行,其结果不能重演,由此,注意到温度在诱导育性转换中的作用。1989年长江流域盛夏低温,引起了许多已稳定的不育系在不育季节的育性波动,从而将温度对育性转换影响的研究提到了切待解决的主要问题。在不同光周期与温度组合的系列条件下,观察光敏核不育系的育性转换,结果见表1:表1 不同光温下农垦58S的育性表现(套袋结实率%) 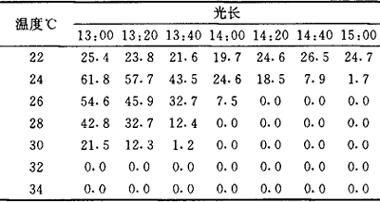
从表中可以看出,光敏核不育系农垦58S不是绝对的光敏不育,其育性转换是受光、温共同作用的结果,当温度高于一定值(32℃)时,在任何光周期下均表现高温不育效应,这一温度称光敏不育性临界上限高温(CST);当温度低于某一值(24℃)时,又在任何光周期下均表现低温可育效应,这一温度称光敏不育性临界下限低温(CFT)。CST至CFT之间(24~32℃)称光敏温度范围(TRPS)。光敏不育系只在TRPS内才表现育性转换的光周期诱导特性。并在TRPS内温度的变化对诱导育性转换的临界光长有补偿作用。即温度升高,临界光长缩短;温度降低,临界光长延长,且对育性转换程度有一定的修饰作用,即短日诱导下的可育度在较高温度下降低;在较低温度下升高。可以认为,在光温敏核不育水稻育性转换的诱导中,光周期及温度的作用是一个连续的数量过程。据研究,由导入农垦58S的光敏不育基因所选育的其它不同遗传背景的不育系,其基因表达的条件,如临界光长、临界温度、光敏温度范围、光温互补作用强弱等,均表现出明显的差异,从而决定了这些不育系的利用价值和适应地区。通常CFT是影响不育系在长日下育性波动的关键因素,CFT高难以保证制种纯度;而CST是影响不育系在短日下繁殖的关键因素,CST低带来不育系在短日高温下繁殖困难。临界光长和光温互补作用强弱是决定不育系适应范围的主要指标。临界光长较长的适宜高纬度地区利用;临界光长较短的适宜低纬度地区利用。光温互补作用强的不育系适应的纬度较宽,在低纬度地区由于温度较高可补偿日照偏短的不足;在高纬度地区则由于日照较长可补偿温度偏低的不足。从实际应用的角度出发,以CST高、CFT低、TRPS宽、光温互补作用强的不育类型最为适宜,但这一理想类型至今尚未见报导。在现有不育系中有一类不育系在长日下不育性过了关,但在短日下繁殖困难;而另一类在短日下繁殖容易,然而在长日下不育性易波动,这是目前两系法杂交稻存在的主要难题。在PGMR育性转换机理方面,虽已证实光敏色素为光受体起着感受光周期的媒介作用,但感受后信息如何控制基因表面尚不清楚,有报导认为PGMR的长日不育与IAA亏缺、GA代谢受阻、ABA过量等有关,另在多胺、醣类、蛋白质及某些酶类等方面,也发现在不同光周期下存在明显或一定的差异。这些工作的积累,无疑对育性转换机理的认识富有启示,但尚缺乏与育性转换直接相关的有力证据。此外,还有报导在不同条件下PGMR幼穗乙烯代谢过旺,且用AVG(ACC合成酶抑制剂)处理,具有导致花粉败育部分逆转的效应,从而启示乙烯可能起着第2信使作用,参与不育基因表达的调控,总之,关于育性转换机理的认识还很肤浅,由于PGMR的育性转换有很大的时空跨度,其机理必然很复杂,尚有待于在光温作用模式下,于雄性器官发育过程中改变其生理生化进程,并结合分子水平上的探索来加以深入研究予以揭示。PGMR的发现为杂交稻利用奠定了改三系法为两系法的种质基础,但要实现这一过渡尚须充分研究育性转换的规律及机理。首先是明确不同遗传背景和农艺性状与PGMR育性转换的光温特性之间的关系,选育出理想类型的不育系。同时应深入探讨其育性转换机理,实现育性转换的人工辅助调控,为某些优良不育系高温影响繁殖或低温影响制种纯度的问题提供技术保障。【参考文献】:1 石明松.对日照长度敏感的隐性雄性不育水稻的发现与初步研究.中国农业科学,1985,(2):44~482 张自国,元生朝,等.光周期条件对湖北光敏核不育水稻育性转换的影响.中国水稻科学,1987,1(3):137~1443 元生朝,张自国,许传桢.光照诱导湖北光敏感核不育水稻育性转变的敏感期及其发育阶段的探讨.作物学报,1988,14(1):7~134 张自国,曾汉来,等.再论光敏核不育水稻的光温作用模式,华中农业大学学报,1992,11(1):1~65 薛光行,赵建宗.水稻光敏感雄性不育临界日长及其对环境因子反应的初步研究.作物学报,1990,16(2):112~1226 孙宗修,等.光敏核不育水到的不光温反应研究.中国水稻科学,1991,5(2):56~607 李合生.HPGMR育性转换与光敏色素相关性的初步研究.华中农业大学学报,1987,6(4):397~3988 徐孟亮,等.HPGMR幼穗发育中IAA的变化.华中农业大学学报,1990,9(4):381~3869 骆炳山,等.光敏核不育水稻育性转换机理初探.华中农业大学学报,1990,1:7~11(华中农业大学张自国撰;骆炳山教授审) |